
Selecting biological control agents
Experimental confirmation of host range
Selecting test species
Every biological control project is unique, and in practice there are no universal rules for selecting which plant or arthropod species should be tested to evaluate the safety of a control agent, or how potential impacts on ecosystems should be assessed. Even without such a fixed framework, applicants to the EPA are required to present a coherent and convincing case that the proposed control agent will not displace or significantly impact on valued flora and fauna. In the past ten years ERMA New Zealand accepted evidence based on as few as two test species (Froud and Stevens 1998) and as many as 46 (see decision on application to ERMA New Zealand NOR02001, go to Documents and Decision [http://www.epa.govt.nz/search-databases/Pages/applications-details.aspx?appID=NOR02001#]) or more. It is up to the applicant to design the risk assessment on knowledge of the biology of the target and its natural enemy, and on ecological and biological control theory.
The strategy proposed by Wapshere (1974) to define the host range of a natural enemy is based on the principle that a plant more closely related to the target weed will present more feeding cues and fewer feeding deterrents to a specialist arthropod herbivore than a plant that is less closely related to the target plants, and will therefore be more likely to be a host for that insect. He formalised this into a procedure called 'centrifugal-phylogenetic method, and this has been the basis for the selection of plants to test ever since. To this he added a set of 'safeguard' criteria for assessing risk to plants that would not be selected on phylogenetic grounds alone, but for which host relationships were not well known. These safeguards were:
- cultivated plants botanically related to the target weed;
- cultivated plants for which there is little or no entomological or mycological knowledge;
- cultivated plants which for evolutionary, geographic or climatic reasons have not been extensively exposed to the candidate agent;
- cultivated plants known to be attacked by organisms closely related to the candidate agent;
- any plants on which the candidate agent has been previously recorded.
Perceptions of risk have changed in the 30 years since Wapshere proposed his protocol. There is greater awareness of the risk posed by colonisation of non-target plants in the receiving range, the possibility of control agents displaying taxonomically 'disjunct' host ranges (Kay and Hill in press), and unacceptable temporary damage caused by 'spill over' feeding (increased awareness of the indirect risks posed by control agents are discussed below).
Briese (2003) suggested three improvements to the criteria for test plant selection that would contribute to better risk analysis:
- use true phylogenetic distance (rather than taxonomic distance) to measure relatedness between plant species;
- consider the degree of biogeographic overlap (this is less relevant for a small landmass like New Zealand, but presumably the same argument can be made for ecoclimatic overlap);
- consider the degree of ecological similarity between potential non-targets.
Despite these recommendations for improvement, it is generally acknowledged that the selection of test plant species to predict the likely host range of weed control agents is well supported by theory.
For oligophagous parasitoids, choosing which species to test, and which factors to incorporate into test designs is more problematic. As it is for weed control agents, the selection of species on the basis of phylogenetic similarity is generally considered a good starting point (van Driesche and Hoddle 1997, Kuhlmann et al. 2005). However, a number of factors make this approach less reliable for parasitoids and predators than it is for weed control agents, including:
- the phylogeny of host arthropods is less well known than the phylogeny of
host plants (
); - most plants are known, but many potential non-target arthropod species are as yet unknown. In New Zealand these may form the bulk of potential species to test (Barratt et al. 2000);
- although known from collection or literature records, some species that should be tested may be unobtainable or impossible to rear (Barratt 2004);
- the host-range of even closely related parasitoids can be disjunct. For many taxonomic groups this pattern is not sufficiently systematic to support an hypothesis as robust as the Wapshere protocols;
- host specificity that appears to be related to taxonomy may instead be related to a correlate of taxonomy, such as the tendency of related species to share habitats (van Driesche and Hoddle 1997);
- many parasitoid groups have predictable host associations but others do not (Sands and Van Driesche 2004). Even amongst predatory arthropods the host range of one species does not reliably mirror the host range of a congener (Causton 2004);
- there are more arthropods from which to choose test plants than there are plants, and so tests of potential hosts of parasitoids are more difficult and expensive than hosts of phytophages;
- it is easier to grow and manipulate plants than insect cultures, and experiments involving insects are more difficult logistically. A list of insects to test must therefore be short;
- rather than being driven largely by phylogenetic similarity, the factors that determine host selection of parasitoids and predators are varied and layered, and difficult to isolate in simple tests (
).
Kuhlmann et al. (2006) described and analysed a range of host-range testing programmes and listed five criteria that have been used to select species for testing against invertebrate biological control agents:
- ecological similarity between host and test species � overlapping geographic range, feeding niche, habitat preference;
- phylogenetic affinity between host and test species - predicted by taxonomic relatedness;
- socio-economic importance of the possible test species� conservation importance, commercial importance, role as a beneficial;
- biological similarity between host and test species � known host range, phenological overlap with the target, dispersal capability, morphological similarity, behavioural similarity, overlap of physiological host range of other agents, response to host plant (Kitt and Keller 1998), similarity of host plant structure (Duan and Messing 1998);
- availability of test species for testing, and the potential for using surrogates.
The information supporting these decisions usually came from the literature, and rarely from first hand experience in the home range of the agent. Babendreier et al. (2005) and Babendreier et al. (2006) also review the criteria for selecting test species. They noted that in many projects only congeners are tested, and recommended improving the margin for error by choosing a wider array of test species.
Recently a decision support system designed to aid the selection of non-target species (NTS) for testing with proposed biological control agents was developed here in New Zealand. The PRONTI (priority ranking of non-target invertebrates) tool was developed for risk assessment biosafety testing of transgenic plants (Todd et al. 2008, Malone et al. 2010, Barratt et al. 2013), and has been adapted and tested for use with entomophagous biological control agents (Todd et al. 2015, Barratt et al. 2016). This was possible because the criteria used to select NTS for biosafety testing with these organisms are fundamentally the same (i.e., the risk posed by the new organism, and the value and testability of the NTS).
Essentially, PRONTI uses a mathematical model and a database of information on NTS, called the Eco Invertebase, to prioritise species for biosafety testing with each proposed biological control agent (Todd et al. 2015). The Eco Invertebase holds information (i.e., taxonomy, food web, biomass, ecology, anthropocentric value and testability data) on more than 1500 invertebrate taxa found in New Zealand. The PRONTI model prioritises the taxa in the Eco Invertebase by applying five selection criteria: (1) the potential hazard posed by the biological control agent to each NTS; (2) the potential degree of exposure of each NTS to the agent; (3) the hypothetical ecological impact that may result from the exposure of the NTS to the hazard posed by the agent; (4) the estimated economic, social and cultural value of each NTS; and (5) the assessed ability to conduct tests with the NTS.
The model output provides information on how each taxon compares with all other taxa in the list using the five criteria. It also provides a measure of certainty of each taxon�s ranking. The model was partially validated by comparing the PRONTI list for the predatory wasp Polistes chinensis with the prey of this species in the field (Todd et al. 2015; see figure below).
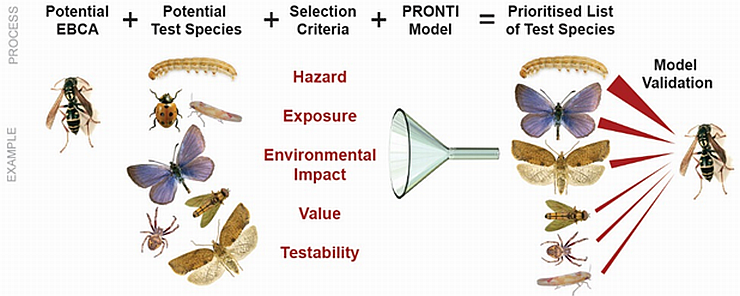
The PRONTI process. The example illustrates the process using the hypothetical biological control agent, the predatory wasp Polistes chinensis, and partial validation of the model through a comparison of the high risk species with the prey items collected from P. chinensis colonies in the field.
© Copyright Plant & Food Research, used with permission.
Currently, PRONTI is undergoing a �live test� with an agent which is proposed for release in New Zealand: Eadya paropsidis Huddleston & Short (Hymenoptera: Braconidae) to target the forestry pest Paropsis charybdis St�l (Coleoptera: Chrysomelidae). Again, PRONTI will be used to rank at least 130 NTS, and the resulting list will be compared with that produced using the currently accepted methodology (Withers et al. 2015). If the PRONTI list is considered to be informative for New Zealand�s Environmental Protection Authority and other decision makers, then PRONTI may become recommended best practice for New Zealand�s biocontrol agent researchers in future. In the meantime if you are interested in testing this tool for your project, contact [email protected]
References
Babendreier D., Bigler F. and Kuhlmann U. (2005). Methods Used to Assess Non-target Effects of Invertebrate Biological Control Agents of Arthropod Pests. BioControl 50: 821-870.
Babendreier D., Bigler F. and Kuhlmann U. (2006). Current status and constraints in the assessment of non-target effects. Pp. 1-13 In: Environmental impact of invertebrates for biological control of arthropods � methods and risk assessment, F. , F. Bigler, D. Babendreier and U. Kuhlmann (Ed.) CABI Publishing, Wallingford, UK
Barratt B.I.P. (2004). Microctonus parasitoids and New Zealand weevils: comparing laboratory estimates of host ranges to realized host ranges. Pp. 103-120 In: Assessing host ranges for parasitoids and predators used for classical biological control: A guide to best practice, R.G. Van Driesche and R. Reardon (Ed.) USDA Forest Service, Morgantown, West Virginia.
Barratt B.I.P., Evans A.A., Ferguson C.M., McNeill M.R. and Addison P. (2000). Phenology of native weevils (Coleoptera: Curculionidae) in New Zealand pastures and parasitism by the introduced braconid, Microctonus aethiopoides Loan (Hymenoptera: Braconidae). New Zealand Journal of Zoology 27: 93-110.
Barratt B.I.P., Todd J. and Malone L.A. (2016). Selecting non-target species for arthropod biological control agent host range testing: evaluation of a novel method. Biological Control 93: 84-92.
Barratt B.I.P., Todd J.H., Ferguson C.M., Crook K., Burgess E.P.J., Barraclough E.I. and Malone L.A. (2013). Biosafety testing of genetically modified ryegrass (Lolium perenne L.) plants using a model for the optimum selection of test invertebrates. Environmental Entomology 42: 820-830
Briese D.T. (2003). The centrifugal phylogenetic method used to select plants for host-specificity testing of weed biological control agents: Can and should it be modernised? In: Improving the selection, testing and evaluation of weed biocontrol agents, H. S. Jacob and D. T. Briese (Ed.) CRC for Australian Weed Management Technical Series no. 7, Adelaide, Australia
Causton C.E. (2004). Predicting the field host range of an introduced predator, Rodolia cardinalis Mulsant, in the Galapagos. Pp. 195-239 In: Assessing host ranges for parasitoids and predators used for classical biological control: A guide to best practice, R. G. Van Driesche and R. Reardon (Ed.) USDA Forest Service, Morgantown, West Virginia
Duan J.J. and Messing R.H. (1998). Effect of Tetrastichus giffardianus (Hymenoptera: Eulophidae) on nontarget flowerhead-feeding tephritids (Diptera: Tephritidae). Biological Control 27: 1022-1028.
Froud K.J. and Stevens P.S., (1998). Parasitism of Heliothrips haemorrhoidalis and two non-target thrips by Thripobius semiluteus (Hymenoptera; Eulophidae) in quarantine. Pp. 526-529 In: Pest Management - Future Challenges. Proceedings of the 6th Australasian Applied Entomological Research Conference, M.P. Zalucki, R.A.I. Drew and G.G. White (Ed.) Brisbane Australia.
Kay N. and Hill R.L. (in press). The disintegration of the Scrophulariaceae and the biological control of Buddleja davidii. In: Proceedings of the XII International Symposium on Biological Control of Weeds, (Ed.) Montpellier, France.
Kitt J.T. and Keller M.A. (1998). Host selection by Aphidius rosae Haliday (Hym., Braconidae) with respect to assessment of host specificity in biological control. Journal of Applied Entomology 122: 57-63
Kuhlmann U., Mason P.G., Hinz H.L., Blossey B., De Clerck-Floate R.A., Dosdall L.M., McCaffrey J.P., Schwarzlaender M., Olfert O., Brodeur J., Gassmann A., McClay A.S. and Wiedenmann R.N. (2006). Avoiding conflicts between insect and weed biological control: selection of non-target species to assess host specificity of cabbage seedpod weevil parasitoids. Journal of Applied Entomology 130: 129-141
Kuhlmann U., Schaffner U. and Mason P.G. (2005). Selection of non-target species for host specificity testing of entomophagous biological control agents. Pp. 566-583 In: Second International Symposium on Biological Control of Arthropods, Davos, Switzerland, 12-16 September, 2005, M.S. Hoddle (Ed.) United States Department of Agriculture, Forest Service, Washington.
Malone L.A., Todd J.H., Burgess E.P.J., Walter C., Wagner A. and Barratt B.I.P. (2010). Developing risk hypotheses and selecting species for assessing non-target impacts of GM trees with novel traits: the case of altered-lignin pine trees. Environmental Biosafety Research 9: 181-198.
Sands D.P.A. and Van Driesche R.G. (2004). Using the scientific literature to estimate the host range of a biological control agent. Pp. 15-23 In: Assessing host ranges for parasitoids and predators used for classical biological control: a guide to best practice, R.G. Van Driesche and R. Reardon (Ed.) USDA Forest Service, Morgantown, West Virginia.
Todd J.H., Barratt B.I.P., Tooman L., Beggs J.R. and Malone L.A. (2015). Selecting non-target species for risk assessment of entomophagous biological control agents: Evaluation of the PRONTI decision-support tool. Biological Control 80: 77-88.
Todd J.H., Ramankutty P., Barraclough E.I. and Malone L.A. (2008). A screening method for prioritizing non-target invertebrates for improved biosafety testing of transgenic crops. Environmental Biosafety Research 7: 35-56.
van Driesche R.G. and Hoddle M. (1997). Should arthropod parasitoids and predators be subject to host range testing when used as biological control agents? Agriculture and Human Values 14: 211-226.
Wapshere A.J. (1974). A strategy for evaluating the safety of organisms for biological weed control. Annals of Applied Biology 77: 201-211.
Withers T.M., Allen G.R. and Reid C.A.M. (2015). Selecting potential non-target species for host range testing of Eadya paropsidis. New Zealand Plant Protection 68: 179-186.
Experimental confirmation of host range | Testing methods |
|---|