
Selecting biological control agents
- Introduction
- Selecting effective agents
- Predicting direct non-target impacts
- Selecting agents for safety
- Exploration in the target home range
- Modelling non-target impacts
- Factors influencing host selection in the target range
- Experimental confirmation of host range
- References
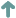
Introduction
This is a selective review of the literature concerning the current state of knowledge about those principles and practices of biological control that are relevant to making an application to the EPA to introduce an exotic biological control agent into New Zealand. It does not cover issues relevant to genetically modified organisms, or to required Maori consultation. It covers the selection and pre-release evaluation of effective and safe agents, be they insects, other arthropods, phytophages, parasitoids, predators, or pathogens of plant and insect pests. It also reviews what we know about assessing the non-target effects of biological control agents on ecosystems and provides some avenues for thinking about these issues. Its purpose is to help applicants to more easily and more completely address the issues that are of importance to the regulator, and hence to environmental, social and cultural biosafety.
The EPA determines whether an agent should be released by weighing the potential benefits of introduction against the potential risks and costs. Applicants can maximise that benefit to cost ratio by:
- selecting the agent or agents best able to reduce the adverse effects of the target;
- knowing how well agent populations will establish, grow and spread;
- identifying what the host range of the agent is likely to be and then testing that hypothesis;
- analysing the likely interactions between the agent and its new ecosystem;
- describing the environmental and ecological benefits of releasing the agent in New Zealand.
The biology and ecology of both targets and potential control agents are hugely diverse, and a 'one-size-fits-all' approach to advice on the selection and development of control agents is not possible (van Driesche 2004). It is unlikely that a prospective applicant will find a perfect model for a proposed biological control project because there is usually too little known and too much diversity. The HSNO process acknowledges that the outcome of a risk/cost/benefit assessment will be uncertain because our knowledge of both the agent and the target are imperfectly known, and because ecosystem responses in the new range cannot be known precisely. The EPA makes decisions in the face of this uncertainty. However, the comfort level of the Authority, and the prospects for a successful application, can be improved considerably by identifying and quantifying those uncertainties, by providing relevant information from parallel projects, and by demonstrating care and attention to the ecological complexity that is inherent in any application. This review facilitates access to theoretical discussions and case studies which will hopefully assist an applicant to consider all relevant facts before applying to import a particular biological control agent.
What is success in biological control?
What the HSNO Act requires
Like the Resource Management Act 1991 [http://www.legislation.govt.nz/act/public/1991/0069/latest/DLM230265.html] before it, the HSNO Act [http://www.legislation.govt.nz/act/public/1996/0030/latest/DLM381222.html] is 'effects-based', enabling legislation. It requires applicants wishing to introduce a new biological control agent to comprehensively assess the probability and magnitude of both the adverse and beneficial effects that might flow from the introduction. The Act is not concerned with changes in the population dynamics, abundance or vigour of a weed or pest resulting from control per se but how those changes affect the pest status of the target. Applicants must focus on the effects of damage to the target rather than the intensity of that damage.
Defining success
Successful control occurs when the abundance or the vigour of the damaging stage falls below a damage threshold (Greathead and Greathead 1992). Rather than the effects of control, percent damage or parasitism of the damaging stage has often been used to describe the impact of biological control agents even though these measures have little relevance to damage threshold, and can sometimes be misleading. For example, the braconid parasitoid Apanteles ruficrus Haliday often kills over 90% of Mythimna separata (Walker) (Noctuidae) larvae in maize crops (Hill 1977, but this level of larval parasitism does not appear to lower the average population level of the pest. In fact, successful control is based more on the reduction in average food consumption as a result of heavy rates of larval parasitism, doubling the tolerance of the plant (Hill 1988). Similarly, very high levels of seed predation by a biological control agent may have little effect on the density of the target weed if the target species is not seed-limited (
). Conversely, life-table studies indicated that only a small increase in generational mortality is required to reduce the population density of codling moth from year to year (Wearing 1979). Immigration of feral females from unsprayed trees play an important role in the population dynamics of the moth, such that even partial success of an efficient biocontrol agent may have significant economic benefits (Wearing and Charles 1989). This illustrates that in intensively managed, high value crops, biological control is but one of many complementary insect pest management tools. It may be difficult to quantify the impact of individual biological control agents, although the sometimes great increase in pests when natural enemies are accidentally killed can dramatically illustrate their collective value. Planning for success
Understanding what is required of biological control should be an important starting point for planning a biological control project, but rarely is. The potential success of a biological control project can only be judged in the context of other management strategies for the pest (
), and defining success at the outset may determine whether biological control is the most appropriate management option at all ( ). Standish et al. (2001) have established a threshold biomass of the forest weed Tradescantia fluminensis below which regeneration of native forest plants is possible. This has become a key performance target in assessing the potential benefits to forest health of a biological control programme against this weed (Hill 2008). Briese (2006) published a protocol used to develop the biological control programme against Onpordum spp. that could equally be applied to the introduction of parasitoids or predators. In most cases where success thresholds for arthropod pests are identified, the research has been retrospective. For example, Barlow and Goldson (1993) showed that the economic threshold for damage to lucerne by the weevil reduction in the density of Sitona discoideus Gyllenhal was 60% of the pest's natural equilibrium population, a level easily achieved by the parasitoid Microctonus aethiopoides Loan. Setting better goals for biological control of both weeds and pests would assist the EPA in evaluating potential benefits.
The approach for weed biocontrol
Cullen (1995) and others have suggested a more systematic approach to predicting the effectiveness of weed control agents, based for the first time not only on specificity of the agent but also on its ability to damage the weed, the environmental factors that might limit agent population density, and the level of damage required to control the weed. More recently there have been calls for population dynamics to be the driving force in agent selection. McEvoy and Coombs (1999) robustly questioned what has been called the 'lottery model' for biological control of weeds - introduction of a number of agents in the expectation that one will work. They considered that introducing multiple agents increased the risk of non-target impacts in North America (however low that might be) without increasing the likelihood of control by any one agent. Instead they also suggested the more parsimonious approach of using modelling approaches to help design biological control systems that targeted the life cycle stages that are both critical to population growth in the target environment and amenable to manipulation.
Sheppard (2003) has made a case for selecting agents in the full knowledge of:
- studies of the ecology and population dynamics of the weeds in the target country, to estimate the key population parameters and life stage transitions as the first step to understanding which factors are most likely to suppress weed populations;
- comparison with the demography of the weed in its native range to help identify the drivers for successful invasion, and the most susceptible life stages of the weed;
- quantitative surveys in the native range that assess the abundance of agents and variation in field impact that can support prioritisation of agents;
- experiments in the native range that manipulate both target and agents to isolate the effects on the population dynamics of the target weed.
Sheppard (2003) points out that advances in control agent selection will only occur when the success of agent selection strategies can be formally assessed retrospectively. This requires setting formal a priori strategies that can be tested over time. This approach is rare (Sheppard 2003), but Briese (2006) provides one such example.
A workshop conducted in Australia in 2004 explored how the effectiveness of biological control of weeds could be improved by the selection of better control agents (Raghu and Van Klinken 2006, van Klinken and Raghu 2006). Using the rangeland weed Parkinsonia aculeata as an example, Raghu et al. (2006) suggested using matrix models in the early stages of programme development to identify the transitions between life stages that were most critical to population growth rate. Having identified the susceptible stages, then modelling or experimentation were used to explore how much change was required in each to achieve a specified population growth level. With these thresholds in mind, the final stage would assess whether each prospective agent was capable of achieving this aim, and (ideally) eliminating from consideration those that were not. The focus of this approach was not minimising the environmental or economic effects of the weed, but on deciding what needed to be done to manage the weed population. However, van Klinken (2006) complemented this study by defining a range of ecological and economic performance criteria (such as reduced patch size, density, and control costs) that would constitute successful management of the weed in pastures or native habitats, and that could be modelled. This approach is closer to the model required by the HSNO Act.
As with the examples provided here, the weed-herbivore models reviewed by Barlow (1999) posed the question 'what degree of control is required to obtain the suppression required' but none explicitly addressed the question of whether agent populations could provide a sustained degree of control.
Morin et al. (2006) proposed a framework for selecting well-adapted pathogens for biocontrol of weeds, critically examined each step, and provided a comprehensive bibliography on pathogen selection. They compared and contrasted their framework with that for selection of herbivorous insects for the same purpose. In the same volume Morin and Edwards (2006) applied this approach to the selection of the most appropriate pathogens for the control of bridal creeper, Asparagus asparagoides.
Testing the host range of an agent is the most costly part of any weed biological control project, and knowing which of the available control agents will be the most effective is likely to save resources and be safer in the long run. Sheppard (2003) concluded that sufficient ecological tools now exist to allow weed researchers to move on from the simple experienced-based value judgements of the past. There is little doubt that ecological studies can assist in better standards for selection of agents, but whether such approaches can be undertaken depends largely on what resources can be accessed for a project, not least the availability of skilled staff in the country of origin of the pest. He acknowledged that even if an ecologically-based approach to selecting agents was preferable, the success and safety rate (Fowler et al. 2000) of what he disparagingly called the 'grab-and-run' approach to biological control of weeds was still high and possibly cost-effective. In fact, most biological control programmes for weeds tend to find a middle course between these extremes. McFadyen (1998) reviews the mixed success of predicting which biological control agents for weeds are best.
The approach for insect biocontrol
Hoddle (2004) noted that the historical success rate for programmes against weeds is three times that for insect pests, and that one reason for this might be the long held requirement for detailed and costly host-range evaluation for weed control agents, leading to a more thoughtful assessment of which agents are likely to work. Certainly, the recent literature suggests that the theoretical basis for selecting biological control agents for weeds outlined here appears better developed than that for invertebrates, although the principles discussed above are equally relevant to the biological control of invertebrates. Barlow (1999) elegantly reviewed the use of 50 models in biological control practice and concluded that improving decision-making by predicting the effect of proposed control agents before introduction was still a largely futuristic goal. The greatest gap in base data appears to be the searching ability, dispersive behaviour and spatial spread of agents, parameters than can only be accurately estimated in the agent's native range. However, he suggested two approaches that could improve agent selection in the absence of such data:
- test a range of scenarios covering the unknown parameters, such as parasitoid searching efficiency;
- using a population dynamics model for the pest, compare the theoretical effects of agents on different life stages of the pest to identify the presence and intensity of density-dependence in the life cycle of the pest. This is analogous to the approach suggested by van Klinken and Raghu (2006) and Briese (2006) for assessing control agents for weeds.
Mills and Getz (1996) also reviewed and critiqued the models available for measuring the efficacy of parasitoids.
Characteristics of successful biological control agents
Perhaps the most successful predictor of how successful an agent will be is its successful use elsewhere. Julien et al. (1999) recorded many examples of the transfer of successful control agents from place to place. The 'Silwood Project' analysed the success or failure of all weed control projects and attempted to draw conclusions about which attributes generally led to control. Conclusions proved difficult, but Crawley (1989) stated that weevils (Coleoptera: Curculionidae) and chrysomelids (Coleoptera: Chrysomelidae) had greater average success rate than other orders. The project also found that characteristics that predicted ability to establish also broadly predicted success, including small size, high voltinism, high fecundity, and long-lived adults. Plant characters that appeared to make plants more susceptible to biological control included genetic uniformity, lack of perennating organs, and susceptibility to secondary infections. Plant characteristics that militated against success included possession of rhizomes, high powers of regrowth and low food quality. Morin et al. (1997) evaluated the relative performance of three agents for mistflower (Ageratina riparia) in Hawaii before selecting a biological control strategy for New Zealand. Froud and Stevens (1998) and Froud and Stevens (2004) selected Thripobius semiluteus as a control agent for glasshouse thrips because it was known to have contributed to control of the thrips in closed habitats in Australia and elsewhere.
The success of related natural enemies and species in the same guild may also help predict success. Conversely, failure elsewhere should contraindicate use in New Zealand, but only if the reason for failure is known. However, this has not stopped the transmission of gorse spider mite worldwide, despite its susceptibility to generalist predators in New Zealand, and wherever it has been released.
Price (2000) contends that bottom-up regulation of insect herbivore populations through plant quality is more common than top-down regulation by natural enemies. Eruptive species tend to be those that feed on a range of plants. This creates the paradox that selecting agents for a high degree of host-specificity also reduces the chance of selecting agents that can outbreak. Stability of the parasitoid-host relationship sometimes reduces control potential.
Mills and Gutierrez (1999) stated that understanding what leads to lasting depression in pest population requires consideration of three processes: spatial heterogeneity, parasitoid coexistence, and tritrophic interactions. Other papers in Hawkins and Cornell (1999) examine the theory surrounding the roles played by ecological factors in the development and implementation of biological control of pests such as spatial heterogeneity and refuges, tritrophic relationships and genetics. Thomas and Waage (1996) demonstrated convincingly how both bottom-up and top-down effects may lead to better pest control than either alone.
McClay and Balciunas (2005) suggested that a pre-release efficacy assessment (PREA) should be conducted as an additional filter in the agent selection process. Although based on weed control agents, their principles of PREA apply equally to arthropod agents. They suggested that predicted impact of an agent could be estimated as:
Impact = Range � Abundance x Per-Capita Impact
Range is estimated by such factors as climatic limits, survival and dispersal, geographic range, diapause and aestivation requirements, and climate matching. Abundance is governed by such factors as voltinism, fecundity, host suitability and survivorship. Per-capita effect can be estimated from native field range studies, experimental manipulation, and from constructs like 'damage curves' - change in fitness with agent load. Many of these contributing factors are discussed in more detail below. In fact, the requirements of the HSNO Act mean that an application to release a control agent could be seen as such an assessment. The selection of effective control agents cannot be concluded in isolation from the effects of land and grazing management (Hatcher and Melander 2003); synergies with other control methods (Buckley et al. 2004); plant competition (for example, Fowler and Griffin 1995, Davies et al. 2005); and more complex tritrophic relationships (Hatcher 1995).
Population matching for maximum performance
Matching agent-host populations
Biocontrol practitioners have generally worked on these two principles: firstly, that the more control agents that attack the pest, the more likely it is that there will be significant reduction in the effects of the pest, and secondly, that agent populations will be greatest when the natural enemy is well adapted to its host and to its new environment. Hufbauer and Roderick (2005) reviewed the importance of microevolution and local adaptation in the success of biological control in detail, and explored whether such generalities were valid. They found examples in the biological control literature illustrating that natural enemies often perform better on genotypes from their local population than genotypes from others, but this was not always true. The strength and importance of local adaptation is influenced by host range, relative generation time, migration rates and the metapopulation structure of host and natural enemy. For example, short-lived natural enemies (such as weed control agents) on long-lived hosts tend to be more locally adapted than on short generation hosts. This may explain the strong emphasis on habitat, biotype and climate matching in the weed biocontrol literature. Similarly, local adaptation is more likely if the migration rate of the host is much lower than that of the natural enemy.
Although not universal, it is clear that local adaptation exists, and genotypic variation of both biological control agents and hosts can influence the level of control achieved. Goolsby et al. (2006) point out examples of where good matching of biological control agent and target has resulted in more host specific and effective agents, and those where agents collected from different biotypes of the host have been ineffective. These authors have reviewed factors to consider during native range research for weed biological control agents. Better awareness of these and other issues reviewed by Hufbauer and Roderick (2005) would allow the selection of better control agents. However, it is clear that occurrence and strength of local adaptation is itself variable from system to system, and theory is not sufficiently well developed to provided day-to-day assistance to practitioners. Clearly these are important issues to address at some level when selecting agents, but the need for more information on local adaptation and its genetics has to be balanced by geographical and financial limitations of overseas research.
Matching agents with the receiving environment
The importance of matching biotypes so that agents can find and exploit the host well is discussed below. The natural enemy must also be physiologically able to survive and reproduce well in its new environment if populations are to build. The climatic conditions in the receiving range must not limit the intrinsic rate of increase of the natural enemy. Both pre-requisites are normally met by introducing a population of the natural enemy that is sourced from the centre of origin of the pest, and from a climate that is broadly similar to the target habitat.
At its simplest, climate-matching must ensure that the temperature and humidity thresholds for survival and development of the introduced agent are met in the new habitat or the agent will not establish. Temperature, humidity and rainfall extremes should not limit survival of any stage. Hill et al. (1993) describe the introduction of additional strains of gorse spider mite to overcome apparent susceptibility to rainfall.
Hufbauer and Roderick (2005) cover a wide range of other issues in their review including the risk that bringing individuals together from disparate locally adapted populations may produce out breeding depression in performance, the distinction and variation between infectivity and virulence (attack and damage) in assessing the value of agents, the role of post-release adaptation in achieving control, the influence of genetic bottlenecks, and the risk of evolution of adverse effects post-release. Increasing awareness of the issues posed by local adaptation is leading more and more to the permitting of only single provenances of agents.
Systems such as CLIMEX and other modelling approaches (
can be used to predict which area within the native range will be an appropriate source of control agents ( and for determining the climate range of both host and natural enemy within the target area. With limited biological information about developmental thresholds and rates, DYMEX modelling can make a first estimate of the life history, potential population increase an distribution of a control agent on release (Kriticos et al. 1999. Barlow (1999) describes several other examples of the use of models to estimate performance post-release.
Conflicts of interest
Apart from issues surrounding unintended and adverse non-target effects, biological control programmes may be challenged because the status of the pest is not universally agreed. This is more likely to be an issue for weed control rather than arthropod control. The biological control programme for gorse (Ulex europaeus L.) began only after consideration of the costs and benefit of the plant indicated that the benefits of reducing the adverse effect of the weed would outweigh the loss of value as deer fodder, as a pollen source for bees, and as a nurse crop for regenerating native forest (Hill and Sandrey 1986). More recently, an application to ERMA (NOC05012 [http://www.erma.govt.nz/search/registers.html?aid=NOC05012]) to introduce biological control agents for broom (Cytisus scoparius (L.) Link) was reconsidered and approved only after a major economic study examined the relative apportioning of costs and benefits to farmers, beekeepers and growers of the fodder tree tagasaste (Jarvis et al. (2006)). Any potential conflicts of interest should be identified before a programme begins in case the conflict is so large that the programme is not viable, or in case long-term data is required to support the eventual case for introduction.
Taxonomic status and synonyms
Clarification of the current taxonomic status is vital to ensure that the correct natural enemy species or entity is introduced. An application to introduce a biological control agent to New Zealand must contain a definitive identification to species level, supported by expert opinion. If this is not possible, evidence must be provided to confirm that the proposed agent is a biologically and reproductively stable entity. For example, Tortrix sensu lato "chrysanthemoides" was approved for release against the weed boneseed in 2004 even though this moth has not yet been formally described (See NOR03001, click on Documents and Decision [http://www.epa.govt.nz/search-databases/Pages/applications-details.aspx?appID=NOR03001#]).
The first step in selecting control agents is to compile information on the identity and taxonomic history of the proposed control agent from literature sources (Sands and Van Driesche 2004), collection data and the expertise of taxonomic specialists. The synonymy may also indicate whether close relatives of the agent occur in the receiving range, providing better definition of likely food web relationships. The current taxonomic status and any past synonyms of the agent must be established before any effort is made to compile the literature of the candidate natural enemy. Unrecognised synonyms can cause important information to be overlooked, and use of an incorrect taxon may lead to the inclusion of false host records (Sands and Van Driesche 2004). The synonymy may provide information on the hosts of related species that might be important in the design of host-range tests. The taxonomic stability of related species may also be important in selecting non-target species for testing. The synonymy of the gorse spider mite, Tetranychus lintearius Dufour included a known pest, which led to experiments to check whether significant hybridisation was likely between gorse spider mite and two pest species (Hill and O'Donnell 1991).
It is equally important to identify the target species. Inconsistent identification of hosts in the native and introduced ranges can lead to the introduction of agents that are either incapable of establishing because the host is absent, or are ill-suited to damage the pest form of the target. Correct matching is most important for highly specific agents such as plant disease pathovars (Morin et al. 2006). The utility of molecular techniques is increasing rapidly, and resolution of these issues will become routine (Sheppard 2003). Better characterisation of population and phylogenetic relationships at the sub-specific level, will result in better physiological and behavioural matches between the agent and host populations, increasing the efficiency of agent selection.
Gassmann and Schroeder (1995) reviewed the major biological control programme against Euphorbia esula L. (leafy spurge) in the U.S.A. After 30 years the taxonomy of the host plant remained difficult, and the taxon was more variable in the native range than in the exotic range. In the native range it grew across a range of biomes and ecoclimatic areas, with various forms, and defining the centre of origin in Eurasia to match US populations was controversial. Although there was a large complex of natural enemies in Europe, they considered that morphological and biochemical differences between the target weed in the native and exotic ranges may have led to host-plant incompatibility, handicapping both the establishment and the performance of the agents released.
First estimation of the host range and relationships: literature
The known host associations of a proposed or selected natural enemy can be accessed from the burgeoning array of electronic and physical information sources now available including abstracts, catalogues, journals, databases, websearch, and other grey literature. Several worldwide catalogues summarise or provide access to host association records:
- arthropods and pathogens for weed control (Julien et al. 1999)
- parasitoids and predators of insects (Thompson and Simmonds 1964-1965, Krombein et al. 1979)
- biological control agents in general including BIOCAT (Greathead and Greathead 1992)
- Review of Applied Entomology
- electronic databases including CAB Abstracts, Agricola, Biosis and Zoological Record
- The internet book of biological control (van Lenteren 2006)
These records are the base data on which the safety of a natural enemy can be initially assessed. However, such information must be interpreted with care. The validity of host records determined from museum specimens and records is limited by the accuracy of the identification of the natural enemy (Sands and van Driesche 2004), and the precision of the host data associated with the specimen. Care must be taken with museum records because of some tendency to 'bulk rear' parasitoids from host insects on field-collected plants. There is a danger then that a parasitoid will emerge from unseen 'other' species, and be attributed to the insect of interest on the plant sample. Many museum specimens of parasitoids are, in fact, collected with no knowledge of their hosts.
Field collection of a phytophage 'from' a plant may not guarantee a host association, since the insect might be simply resting on the plant. The enshrining of dubious records in databases may lead to overestimation of the host-range, and such records should not automatically exclude a potential agent from further assessment. Host range can usually only be determined by rearing them from a host. Some host records may originate from laboratory studies, and may reflect the physiological host range rather than the likely range of the natural enemy in the field. For well-known control agents such as Macrocentrus grandii (Braconidae), gaps in knowledge can in themselves be useful indicators of host-range (De Nardo and Hopper 2004).
Comparisons of associations in native and receiving range
Apart from the records of host associations in the native range of the natural enemy, other records essential to the assessment of both the efficacy and risk of a biological control agent are:
- other hosts of the proposed control agent in the native range;
- other hosts of the control agent where it has established outside its native range;
- other natural enemies of the host in the native range;
- other natural enemies of the host where it has established outside its native range;
- natural enemies of hosts in the native range that are related to the target (Hoddle 2004);
- natural enemies of hosts in the exotic range that are related to the target;
- natural enemies of hosts related to the target in the receiving range (Hill and Hulley 1995).
For all the wealth of information now available, it is almost certain that there will be insufficient recorded information to adequately define the risks posed by the proposed natural enemy to non-target species and ecosystems. The non-target fauna in the receiving region may be under-studied or the agents may even be new to science. Gaps in the identification of fauna at risk in the receiving range can be filled by developing a list of species at risk based on critically examining the phylogeny, morphology, physiology, behaviour, geography, phenology, vagility, climatic requirements and habitat preferences or requirements of both agents and potential hosts (Hoddle 2004).
Lack of host records where they might be otherwise expected may be useful in identifying primary host range or ecological limits to host use (Sands and Van Driesche 2004).
Sources of information about host associations for natural enemies in New Zealand include:
- the plant and insect databases at
Landcare Research [http://www.landcareresearch.co.nz/databases];
- Te Papa Tongarewa and other museums and institutions;
- the BUGS database;
- PlantSyNZ database [http://dataversity.org.nz/guide/systems/plant_synz/];
- PIPN database (MPI Biosecurity);
- Forest Research Institute collection
Realising the value of New Zealand's databases for biological control
New Zealand's agricultural landscape is remarkable for the fact that it is composed almost entirely of Northern Hemisphere plant taxa and is essentially no more than 150 years old. The majority of species in these new agricultural landscapes are also exotic invaders from the North, although a few intrepid native species have also successfully adapted to these novel landscapes. The exotic invaders include both pests and natural enemies, and the latter include suites of parasitoids and predators, most of which arrived accidentally over time rather than as part of deliberate biological control programmes (Charles 1998).
New Zealand's long history of activity in biological control and ecological research in agricultural ecosystems (Cameron et al. 1989) means that there is an extensive database of many of these species (Ferguson et al. 2007). Some have been here for many decades, others continue to arrive annually at present. Those that have been here for some time provide a valuable resource for risk analysis, in that it is possible to measure the actual, realised extent of their impact on native fauna and compare that with criteria that would be considered or predicted by modern host-range testing in quarantine conditions. This approach can allow direct testing of some of the bullet points posed above (e.g. Charles and Allan 2002 and ongoing research). If and where ecological 'safety' of related natural enemies can be demonstrated, then such empirical data can add confidence to host-specificity predictions of proposed new, biocontrol agents. Improved knowledge of the ecology of New Zealand's exotic natural enemy fauna may hence provide a valuable 'anchor' for future biocontrol theory, host-range testing procedures, and new biocontrol programmes.
Natural host range
The host range of a biological control agent in the area of origin is potentially useful as a guide to likely host range in new areas of introduction. If available, this information can assist in risk assessment and prediction of post-release impacts. Regulators considering applications for the release of new biological control agents often request information about host range in the country of origin and in other areas where introductions have occurred, for example, the FAO 'Code of Conduct' for import and release of biological control agents (FAO 1996). Aquiring information about natural host range is considered to be an important first step in the process of biological control agent host specificity testing, particularly selection of test species (Kuhlmann et al. 2005). This can help substantiate the interpretation of host range tests in containment, and provide support to the hypothesis that host range tests in containment have over-estimated field host range, if data from the natural range indicate that a particular species has not been attacked in the field. Several studies have shown that fundamental host range is greater than ecological host range (e.g. Cameron and Walker 1997). Furthermore, a comparison of the value of laboratory tests as a predictor of ecological host range in the area of origin with a proposed area of release might also be of assistance in interpreting containment host range data.
The approach to this work is likely to be different for weed compared with insect biological control agents. In some ways initial information for the latter is more easily achieved because while collecting specimens of the target to rear out parasitoids, or culture pathogens, other species are likely to be collected from the same habitat. The challenge then becomes rearing a number of different species, although for parasitoids, dissections can be carried out to determine parasitism. For weed biological control agents, it is necessary either to observe impacts on other plants and determine whether the candidate biological control agent was responsible, or to carry out laboratory or field tests confining the proposed agent on a range of plants.
During the exploration phase of a biological control programme, target host populations are naturally the focus of investigation, and related or co-existing species are often not examined, especially in the case of entomophagous species. Weed biocontrol programmes do sometimes include examination of other plants in the area, and plants which are phylogenetically related.
While there are many examples of this approach having been taken during biological control programmes, few studies have been undertaken either retrospectively or post-release to test the accuracy of predictions made from native and novel host range studies. Those that have been carried out, show in general that research conducted in the natural geographical range of a proposed biological control agent has or could have assisted in predicting the likely breadth of post-release host range of biological control agents. Some examples are given in the next section.
Some case studies
The natural host range of Coleoptera feeding on broom (Cytisus scoparius) was surveyed in Europe by Syrett and Emberson (1997) as part of a biocontrol programme for broom. Their aim was to obtain field data on host preferences of broom-feeding Coleoptera to assist in the selection of the most host-specific species for a biocontrol programme. They found nine species feeding on species in the same genus, but that two, Bruchidius villosus (F.)(Coleoptera: Chrysomelidae) and Exapion elongatissimum (Desbrochers)(Coleoptera: Brentidae) were considered to be essentially monophagous. Subsequently, after release in New Zealand it was found that B. villosus attacked the exotic non-target species tagasaste (Cytisus proliferus). Further studies in the native range showed that B. villosus aggregates on the earliest-flowering species, which in New Zealand is tagasaste, which flowers before broom (Sheppard et al. 2006). This study emphasised the value of native-range studies in understanding the role of phenological synchrony in ecological host-specificity.
Haye et al. (2005) retrospectively determined the ecological host range of the braconid Peristenus digoneutis Loan introduced into North America from Europe for control of Lygus spp. (Hemiptera: Miridae). Potential non-target mirids were collected from a wide range of environments in northern Germany. They found that the parasitoid was reared from seven non-target hosts in the same subfamily as the target host but no other subfamilies, and that overall parasitism in non-target species was never more than 1%. No-choice laboratory tests in North America suggested that native Lygus spp. were likely to be hosts but other non-target Miridae were not suitable hosts for P. digoneutis (Mason et al. 2011). Post-release studies in North America have shown that three non-target mirid species were attacked, but at very low levels. However, Peristenus relictus (Ruthe), another parasitoid of mirids was predicted to attack several other non-target species in the field (Mason et al. 2011). The authors noted that although a number of non-target hosts have been identified in the area of origin, and area of introduction, severe non-target effects are not necessarily indicated. They concluded that a complete host range evaluation should combine data from field collections in the area of origin, and laboratory data to avoid rejecting promising biological control agents.
A study of the natural host range of the tachinid Aphantorhaphopsis samarensis (Villeneuve) being considered for biocontrol of gypsy moth (Lymantria dispar L.) was carried out. The only known hosts from the literature were the lymantriids L. dispar and Orgyia recens (Hubner) (Fuester et al. 2001). Collections of potential hosts in Europe in areas where A. samarensis was abundant produced no records of additional hosts, or where pupae were found that resembled the parasitoid, no adult successfully emerged. Over a five year period about 850 larvae representing 54 species and 11 families of Lepidoptera, there were no records of parasitism by A. samarensis. Host specificity testing with North American Lepidoptera were consistent with the data obtained from Europe, and it was concluded that A. samarensis has a narrow host range and presents minimal threat to non-target species.
A review of the research carried out in the natural range of Rhinocyllus conicus (Fr�l.) was carried out to determine whether the non-target impacts observed in the introduced range in North America on native thistles could have been predicted (Gassman and Louda 2001). Surveys in Europe over the period 1961 to 1982 showed that R. conicus larvae were found on species in the genera Carduus, Cirsium, Silybum and Onopordum, although a preference for Carduus was indicated. Initially differential phenology was thought to be responsible for geographical variability in host range, but later biotype differences in R. conicus were implicated, and a further theory about innate preference resulting from resource concentration was put forward. The latter two factors combined to convince biocontrol practitioners that there would be minimal non-target impact on native Cirsium species in North America. However, comprehensive post-release studies have shown that R. conicus has undoubtedly impacted on native Cirsium populations in North America. The authors suggest that the evidence from the natural range was sufficient to indicate that further tests were required before a decision to import R. conicus should have been made, and a full analysis of the scientific evidence was sufficient to predict that non-target impacts would occur. They concluded that pressure to address the weed problem, focus on non-target effects on economic plants, and the biotype argument led to the decision that was made.
Gassmann et al. (2008) carried out field surveys in Europe to identify herbivorous arthropods which were genus-specific to the buckthorn species Rhamnus cathartica and Frangula alnus for biocontrol in North America. They identified 21 genus-specific herbivores and 17 apparently monophagous species amongst the herbivorous fauna of buckthorn, and discussed the feeding strategies of these in relation to selecting potential biological control agents.
In New Zealand the braconid Microctonus aethiopoides Loan was introduced to control Sitona discoideus Gyllenhal, but post-release studies have shown that nine genera of non-target species are attacked in the field (
), and retrospective laboratory host range tests have shown that nine genera were attacked (five in common). The same parasitoid was introduced in to Australia, and surveys to date have shown that only one native species (single record) was attacked (Barratt et al. 2005). M. aethiopoides was introduced from Moroccan populations to both Australia and New Zealand, but the known natural host range includes only two genera of weevils, Sitona and Hypera. Recent investigations in the natural range, Morocco, and in Australia where the previous record of non-target parasitism was found, were carried out to determine whether M. aethiopoides is indeed restricted to these two genera in Morocco, or whether in fact the natural host range is much broader. Over 600 weevils comprising 47 species were collected in Morocco, but only four species were found to contain parasitoid larvae consistent with M. aethiopoides. These were Sitona discoideus, which was the species most abundantly collected, but also parasitised was Charagmus gressorius (F.), C. griseus and Hypera postica (Gyllenhal).
Sampling in Australia was also designed to optimise collection of potential non-target hosts of M. aethiopoides. However, no further records of non-target parasitism were detected after collecting about 350 specimens of weevils comprising 13 species from and near lucerne at the site in NSW where the single incidence of non-target parasitism had been found previously.
Modelling non-target impacts
The focus of this section is on evaluating whether models can help predict the impacts of a candidate biological control agent (BCA) on its target and nontarget hosts prior to release. Note that this also has direct application to the broader issue of predicting the risks posed by any new species, whether deliberately or accidentally introduced.
A particular challenge for models is to make effective use of the limited information that is typically available before an introduction. In most cases we will know the life cycle and offshore distribution of the candidate BCA. Pre-release testing is likely to provide indications of host range in the proposed area of introduction, attack rates, and development rates at particular temperatures. The literature may provide further information, but for the purpose of developing general approaches to predicting impacts, the models must be able to be parameterised from these limited data.
A vision for the future is that a suite of validated modelling approaches will be available for predicting the impacts of candidate BCAs on resident species. The models will utilise different levels of biological detail so that some insight can be derived from the most minimal data, with greater accuracy and precision being possible with increasing biological knowledge. Some information on resident species is needed, but the models will help to target the collection of relevant data and utilise it for different introductions. Ultimately, the models will allow more robust decisions to be made about deliberate introductions and their likely risks and benefits.
This section briefly reviews the models and approaches that have been developed to date, and describes how they could be further progressed to better predict the impacts of deliberate introductions.
Attack rate and population impact
The success of a BCA is usually estimated by measuring the proportion of the pest or weed population that is attacked in the field. Similarly, nontarget impacts are usually quantified in terms of the proportion of nontarget individuals attacked (Mills and Kean 2010). However, this is misleading since host population size may or may not be significantly affected by a BCA, irrespective of the proportion attacked (Barratt et al. 2010). Relative population size with and without BCA attack will almost always be the most relevant measure of BCA impact, so the relationship between attack rate and population size is fundamental to assessing impact.
Kean and Barlow (2000) and Barlow et al. (2004) were the first to use models to explicitly explore the relationship between BCA attack rate and host population suppression, in the contexts of target and non-target impacts, respectively. These models have been advanced by developing an overarching framework which encompasses any enemy-host life cycle, relates the previous models to each other, and suggests a hypothesis to explain the empirical relationship between maximum attack rate and the probability of successful biological control noted by Hawkins and Cornell (1994).
Estimation of population impact from a given attack rate, and the attack rate leading to host extinction, requires knowledge of the host life cycle and intrinsic rate of increase; these are basic parameters that may be estimated relatively easily. For deliberate introductions, indicative attack rates may be estimated from pre-release host range testing experiments. Alternatively, the models are simple enough that decision-makers can easily simulate "what if?" scenarios to explore the implications of uncertainty in the parameters. An interactive web page is available for this purpose.
Relationship between attack rate and impact for low and high altitude native weevil populations in Otago.
The figure shows an application of this model to native weevils under nontarget attack by the parasitoid Microctonus aethiopoides in Otago tussock grasslands. The intrinsic rate of increase of these weevil populations declines with altitude (Barlow et al. 2004). While neither upland nor lowland populations are likely to be significantly reduced by the 2% average attack rate observed (dotted line), both would be substantially reduced by the 24% maximum attack rate observed (Barratt et al. 2007), should this be sustained (dashed line). Under the latter scenario, the model suggests that lowland populations would be reduced to 64% of their natural density and upland populations to 24%. The model also suggests that lowland populations will be driven extinct by sustained attack rates of greater than 53% and upland populations by greater than 30%. The ability to estimate population impact from attack rate will be of enormous benefit to the EPA's and DOC's reviews of proposals for future proposed BCA introductions.
Future research should emphasise validation of the models using meta-analysis of published results from the literature, and post-release studies of biological control introductions such as those being conducted in projects being carried out in the research programme Better Border Biosecurity [http://www.b3nz.org].
Interactive model for predicting host impact from natural enemy attack rate
This tool allows the relationship between attack rate and population impact to be explored interactively.
Note that these models assume discrete host generations.
Reference: Barlow et al. (2004)
Primary versus secondary attack
In the context of predicting nontarget impacts from candidate BCAs, an important issue is whether the attacked species is a primary or secondary host. Primary hosts are those that can support the BCA even when no other hosts are available, while secondary hosts are those that cannot support the BCA in the absence of other host species. Therefore, impacts on secondary hosts arise largely from "spill-over" attack from primary hosts, and removal of spill-over sources will allow recovery of secondary hosts. BCA population growth will be constrained by the availability of primary hosts, but will be independent of the density of secondary hosts. Therefore, secondary hosts could suffer rates of attack that are sufficient to cause local extinction, but this is unlikely to occur in primary hosts because BCA populations will decline in tandem with reductions in their primary host populations.
Previously published models have been analysed to elucidate the differing drivers for population impact in primary and secondary hosts, and composite models have been developed to explore intermediate scenarios. In addition, these ideas have been applied in the context of nontarget parasitism of native weevils by the Moroccan biotype of Microctonus aethiopoides in mid-altitude tussock grasslands in Otago. M. aethiopoides has been observed to parasitise up to 24% (mean = 2%) of native weevils in the vicinity of populations of the target host, Sitona discoideus (Barratt et al. 2007), but it is not known whether these are primary or secondary hosts. M. aethiopoides can develop through multiple generations on the non-target native weevil Niceana cervina in the laboratory McNeill et al. 2009, but experimental releases of the parasitoid on the Lammermoor-Rock and Pillar Range failed to augment parasitism rates in native weevils.

Moroccan Microctonus aethiopoides on Sitona discoideus.
© Copyright AgResearch, used with permission.
The ability of the target host S. discoideus to reproduce on host plants other than its preferred host, lucerne, is confirmed by its establishment on Norfolk Island where lucerne is absent (Vink and Phillips 2007). In the laboratory, M. aethiopoides survives and develops equally well in S. discoideus adults fed on clover (present as a weed in tussock grasslands) as in those fed on lucerne (absent from tussock grasslands), suggesting a pathway for nontarget parasitism via spill-over immigration of S. discoideus to tussock grasslands. In addition, the potential for S. discoideus to survive on clover in these environments raises the possibility of resident S. discoideus populations acting as a local source of nontarget parasitism. However, it is not yet known whether such resident S. discoideus populations do persist in native tussock grasslands. An ad hoc survey failed to detect and S. discoideus larvae on the roots of white clover on the Lammermoor-Rock and Pillar Range (B. Barratt, pers. comm.). In addition, field experiments have shown heavy mortality of M. aethiopoides pupae and parasitised native weevils over the winter, but sufficient survival of the latter to allow a few parasitoid adults to emerge in spring. On current evidence, it seems most likely that the primary source of nontarget parasitism by M. aethiopoides in mid-altitude tussock grasslands is spill-over from the target host, perhaps slightly augmented by local population cycling within native weevils.
The modelling analyses of the impacts of BCAs on their primary and secondary hosts are improving our ability to define risks to nontarget species, but further development and validation is required. The role of spill-over effects is often addressed in applications to the EPA, but these are usually hypothetical with no supporting data or analytical framework. The modelling work described above has potential both to provide the needed analytical framework, and to clarify the types of supporting data that applicants must present to the EPA to ensure robust decisions can be made.
Climate matching
Climate matching has been widely used to predict the potential distributions of introduced plants and arthropods under current and future climates (Kriticos and Randall 2001). For example, CLIMEX software (Sutherst and Maywald 1985, Sutherst et al. 2007) empirically fits an eco-climatic envelope to the native distribution of a species and uses this to predict its potential introduced range. However, it is not clear how the relative ecoclimatic suitabilities of an area for a BCA and its target and nontarget hosts may affect the likelihood of impacts on each host. In the absence of data, some preliminary hypotheses were erected and explored using the case study system of an introduced BCA, the herbivorous weevil Cleopus japonicus, its target host plant Buddleja davidii and a known nontarget host plant Scrophularia auriculata. The hypotheses were: (1) impacts of the BCA on the target host plant are greatest in areas which are climatically optimal for the BCA but marginal for the plant, and (2) impacts of the BCA on the nontarget plant are greatest in areas which are marginal both for the target plant and the nontarget plant, but optimal for the BCA.
CLIMEX models were developed for C. japonicus and B. davidii in New Zealand, and the intent was to set up field experiments in localities with different climates to test the above hypotheses. However, the predicted potential distributions for both species were found to be very similar, making this a poor system for testing the hypotheses. Further work using other case studies may advance these ideas and eventually lead to a new tool for identifying where target and nontarget impacts are likely to be the greatest. This knowledge would help to design release, monitoring and management programmes for future BCA introductions.
Phenology models
Phenology models often use the thermal requirements for development of an insect to predict the duration and timing of life stages in a novel environment (Barlow et al. 1994, Kean and Kumarasinghe 2007). Several phenology modelling approaches have been advanced to inform the risk of species introductions. Circle map analysis (Gurney et al. 1992, Powell and Logan 2005) was used to assess the ability of the parasitoid Microctonus aethiopoides to survive and persist in nontarget native weevil hosts in Otago. Logged air and litter temperatures from three tussock grassland sites were used together with parasitoid development rates (Goldson et al. 1990) to show that 2-3 parasitoid generations were possible and that resident parasitoids were unlikely to be present as potentially cold-susceptible pupae and adults in winter. However, depending on the timing of introduction, survival of first generation immigrants may be unlikely. Nevertheless, the models suggest that there are no major temperature constraints on the ability of M. aethiopoides to parasitise nontarget weevils in mid-altitude tussock grasslands. This contributes to the understanding of this system, which has been an influential case study for understanding nontarget effects of BCAs.
Another use for phenology modelling in estimating likely impacts is to examine the generation time of a natural enemy relative to that of its host (generation time ratio, GTR). It has been shown that for aphids and their predators GTR is a strong predictor of host suppression (Kindlmann and Dixon 1999). The theory has also been applied to parasitoid-host systems (Mills 2006), and the success and failure of some classical biological control introductions to New Zealand have been assessed in terms of their GTR (Barlow et al. 2002). Preliminary work has shown that the thermal development and diapause requirements of BCAs and their hosts can be used to estimate the GTRs on a spatially-explicit basis, potentially identifying areas of high and low likely impacts. Further work is needed to validate the GTR hypothesis in this context.

Microctonus hyperodae parasitising the Argentine stem weevil, Listronotus bonariensis.
© Copyright AgResearch, used with permission.
Phenology models may also be used to assess the likely synchrony of BCAs with their potential hosts. This has particular potential for identifying periods of high risk of nontarget attack by BCAs. For example, Barlow et al. (1994) developed a phenological model to help plan and monitor the classical biological control introduction of the parasitoid Microctonus hyperodae against the Argentine stem weevil, Listronotus bonariensis, in New Zealand. The model also suggested that parasitoid adults would be likely to peak in spring at a time when relatively few adult weevils would be available for attack, and this has subsequently been shown to occur in the field (Goldson et al. 1998, Phillips et al. 1998). Historical records of nontarget pasture weevils have been used to map the seasonal abundance of these species in relation to M. hyperodae and L. bonariensis. This suggests that the risk of nontarget attack during the spring may be relatively high, especially for the more abundant species at this time, Steriphus variabilis in Canterbury and Irenimus aequalis in the Waikato. Indeed, low levels of parasitism of these species by M. hyperodae have been observed at these sites (Barratt et al. 1997, Barratt et al. 2000). This work demonstrates the potential for phenology models for introduced species, together with knowledge of resident species' populations, to predict when nontarget attacks might occur, and thereby guide monitoring and management.
Detailed population models
A deliberate introduction for the purpose of classical biological control typically involves a programme of a priori research to estimate the risks and benefits of the introduction, and this presents an opportunity to collect the data needed to parameterize more detailed models for the impacts on target and nontarget species. For example, Raghu et al. (2007) used pre-introduction native range and quarantine experiments to parameterise a model for the likely impacts of a herbivorous beetle on target and nontarget plants in Australia.

Cleopus japonicus on Buddleja davidii.
© Copyright Scion, used with permission.
A similar modelling approach was used to predict the impacts of the herbivorous weevil Cleopus japonicus on Buddleja davidii and nontarget plants in New Zealand. A detailed model was developed, using climate variables to drive weevil development and plant growth rates, and to predict when food shortages would drive C. japonicus to spill over to nontarget plants. The model suggested that emigration to nontarget host plants was likely to occur in late summer or autumn, with the magnitude of the nontarget threat being greatest at relatively warm sites. These predictions were borne out by the field trials at four sites (Watson 2007), in which spill-over damage to Scrophularia auriculata occurred in March when defoliation of B. davidii was greatest. Significantly, S. auriculata is an annual plant which sets its seed in summer, so defoliation in March is unlikely to have any impact on its population density. This work showed that detailed population models can successfully predict the timing and impact of nontarget attack from introduced species.
What can be predicted?
These modelling approaches predict three aspects of the potential nontarget impacts of an introduced BCA: (1) the likely affect on nontarget population size or density from a given attack rate by an introduced BCA; (2) the likely seasonal timing of new BCA attack and how this may affect its impacts on nontarget species; and (3) the geographic locations where impacts are likely to be the greatest (see table). These approaches can also be applied to predicting the impacts of accidental introductions.
Characteristics of the impacts of a biological introduction that may be predicted by different modeling approaches. Brackets indicate areas where further research is required to demonstrate potential. Characteristic
Attack rate
and impactClimate
matchingPhenology
modelsDetailed
population modelsMagnitude of local impacts 
( )( )
Seasonal timing of impacts
Geographical location of greatest impacts ( )( )
The availability of suitable data is probably the biggest constraint to using models for predicting the nontarget impacts of biological control introductions. The following table summarises some potential sources of data for parameterising the models discussed above.
Potential sources of data for parameterising models to predict the impacts of biological introductions. Brackets indicate where particular care should be taken in applying the data available. Sources of data
Attack rate
and impactClimate
matchingPhenology
modelsDetailed
population modelsScientific literature
Herbarium/collection records ( )Pre-release host range testing
Other quarantine studies ( )
Ecological studies from the native/exotic range
Extrapolation from closely related taxa ( )
How can models improve the biosafety of future introductions?
Minimum Standards (section 36 of the HSNO Act 1996) must be met for the EPA to approve an application. Grounds for declining an application include displacement of any native species within its natural habitat, significant deterioration of natural habitats, or any adverse effect on New Zealand's inherent genetic diversity. The applicant's risk analysis must address these issues, but its quality is severely constrained by limitations both of testing organisms in containment, and in the information that can be derived from the receiving environment before release. The models discussed in this section can help applicants, the EPA, and other stakeholders to predict when, where and how much impact will arise from a proposed biological control agent introduction. In addition, the models highlight the biological parameters that have the greatest influence on impact, thereby identifying the most important data to obtain. Application of the analytical frameworks provided by predictive models can add considerable value to the applicant's risk/benefit analysis.
Models can also help to optimise post-release monitoring by indicating which nontarget species should be monitored, what time of year they should be examined, and what geographic regions should be sampled. Retrospective post-release analysis of impacts is the only way for regulators to validate their risk analysis methods and decision making processes. As demonstrated above, models can also assist in understanding post-release validation case studies. Well structured and analysed information on previous case studies adds significant value to the decision-making process by providing insights to likely outcomes from future introductions.
Validation of the models is a priority for future research, and there is significant potential to capitalise on other research projects for this. In particular, there is considerable scope to learn from deliberate introductions to improve models for predicting the impacts of accidentally introduced species.
Geographic range and biotypes
As mentioned earlier, host range and other biological characteristics of organisms may vary from one population to another across their native range. Where these differences can be defined the population can be described as a strain or biotype: a group of organisms of the same species that have clear-cut physiological but not necessarily morphological distinctions from others within the species. Biotypes can normally interbreed, although biotypes can have sexual and parthenogenetic forms (Gerard et al. 2006). The existence of biotypes can often be inferred from the diversity of host records that appear in the literature. The Thompson and Simmonds catalogue (Thompson and Simmonds 1964-1965) records a wider host range for many oligophagous parasitoids than can be measured in any one population (Hill et al. 1985).
The existence of previously unrecognised biotypes is gradually becoming much better understood. The target or the natural enemy may actually consist of several distinct populations that vary biologically but are lumped under a single taxonomic name. The existence of variation between populations of natural enemies creates both risks and opportunities in the development of a biological control programme. For example, unless biotypes are distinguished, a complex of narrowly specific parasitoids might be incorrectly viewed as one widespread, more polyphagous species (Sands and Van Driesche 2004), risking the rejection of potentially useful agents. Selecting the wrong source population can compromise the probability of establishment or successful control of the target pest.
Biotrophic plant pathogens such as rust fungi can be particularly specific to different forms of target weeds. Puccinia chondrillina has successfully controlled the narrow-leafed form of skeleton weed, Chondrilla juncea, but after over 30 years has not adapted to attack the other forms. Similarly, the biological control programme for blackberry in Australia has involved the selection of a range of pathotypes of Phragmidium violaceum that are specific to different groups of the Rubus fruticosus aggregate (Morin et al. 2006).
On the other hand, the inclusion of genotypes from a population that has not been subjected to host range tests can lead to unpredicted host use following release (Quentin Paynter, pers. comm.). This was the case when the gorse pod moth, Cydia succedana, was introduced to New Zealand from the UK in 1992 as a biological control agent for gorse. This species now makes occasional use of non-target plants at levels that were not predicted by host-range testing (Paynter et al. 2004). This has been attributed to the release of untested genotypes from Portugal that have a subtly different pattern of host use (Quentin Paynter, pers. comm.). It is now standard practice in New Zealand to release only those populations of weed control agents for which host range tests results exist. Sands and Van Driesche (2004) recommended that host range tests for parasitoids should be also be conducted using the population that is to be released to avoid the possibility of using different biotypes which might have different host ranges.
Variation between populations has provided a potentially powerful tool for biological control of Sitona lepidus in New Zealand. Microctonus aethiopoides Loan was introduced to New Zealand in 1982, and provides adequate control of Sitona discoideus Gyllenhal. When clover root weevil (S. lepidus Gyllenhal) became established, the New Zealand resident M. aethiopoides failed to parasitise the new pest, even though this was a common parasitoid of S. lepidus in its native range (Barratt et al. 1997). Before a new strain of the parasitoid could be introduced from Europe, rearing experiments indicated that interbreeding between the new S. lepidus strain and the resident S. discoideus strain led to poor performance in hybrids, and the prospect of failure in the control of S. discoideus should the new biotype be introduced (Goldson et al. 2003). Instead, a parthenogenetic strain, incapable of interbreeding with the biotype already in New Zealand, has been introduced (Gerard et al. 2006). These forms of the parasitoid can now be distinguished using molecular techniques (Vink et al. 2003) and there may be cryptic species involved. Goldson et al. (1997) also discuss the potential of differing parasitoid strains in biological control.
For many years the best-adapted control agents were thought to occur at the 'centre of origin' of a pest. The competing theory that the most dynamic control agents would be 'new associations' between natural enemies and unusual hosts, often from the edge of the native range of a pest (Hokkanen and Pimentel 1989) was considered particularly high-risk by Simberloff and Stiling (1996).
Fundamental vs ecological host range
Fundamental host range of a natural enemy is defined as the sum of all species on which the agent can complete its life cycle, and the sole barrier to host use is whether the host stage is an acceptable substrate for development. This might more accurately be described as the taxonomic rather than the phylogenetic host range, because nomenclature does not always reflect phylogeny.
The ecological or realised host range is the subset of the fundamental host range that can be used by the natural enemy under natural conditions where there are additional, ecological barriers to host use. Not only must the host be suitable to support development of the immature control agent, but females must be able to locate a susceptible host and lay fertile eggs there. Some of the ecological and behavioural barriers that must be overcome to ensure this include host-acceptance cues and host-finding, niche-finding, habitat preference and life cycle synchrony. These factors all affect host use and should be considered in assessing the risk of non-target attack.
For biological control of weeds it is generally accepted that simplistic small arena tests estimate fundamental host range well but overestimate the ecological host range because host acceptance/rejection mechanisms are compromised (Cullen 1989). The behaviours leading to host acceptance or rejection in parasitoids are often sequential and more complex, and often are not independent of the substrate and prior experience of the agent. Small arena tests may therefore overestimate or underestimate the host range depending on the parasitoid/host system under study. Vinson (1998) summarised the mechanisms leading to successful parasitism as:
- host habitat location;
- host location;
- host acceptance;
- examination;
- probing;
- drilling;
- oviposition;
- suitability.
Host suitability and physiology
The nature of the physiological relationship between the control agent and its host ultimately controls the degree of host specificity, and the most categorical measure is whether the host can support complete development of the agent or not. There is considerable evidence to back the hypothesis that herbivorous insects (and diseases) have evolved to exploit particular plant species share common features of chemistry, form, and/or phenology. Secondary plant chemistry usually closely tracks host taxonomy. Although the relationship is not nearly so clear cut, this is also true for parasitoids and predators (van Driesche 2004, Kuhlmann et al. 2005). Strict monophagy is not uncommon amongst parasitoids, and the ever growing body of observation shows that specialisations are often taxonomically conserved within groupings (Kuhlmann et al. 2005. There are many examples: for instance we know that aphidiine braconids only attack aphids and that Metaphycus species only attack soft scales in the family Coccidae (Sands and Van Driesche 2004). This is not surprising as closely related hosts will share similar physiology and defensive mechanisms, and will tend to occupy similar habitats. However, we also know that host ranges are often disjunct between genera, and the factors that mediate host range in parasitoids and predators do not appear to be as simple or straightforward as those for weed biocontrol.
Larval survival in the host may be the most discriminatory stage in determining the host range of a parasitoid (van Driesche and Murray 2004). Those that cause their hosts to cease development once parasitised are called idiobionts, and are usually external or egg parasitoids. Koinobionts live within the host and manipulate and overcome the immune response to maintain a favourable environment in which to grow. The complex mechanisms by which koinobiont larvae influence host defences include venoms and toxins, and even symbiosis with polydnaviruses (
). Parasitoids that contend with physiological host defences in this way probably require very specific adaptations to the physiology of the host and will tend to be host-specific. Idiobionts do not require such finely-tuned physiological adaptation and tend to be more polyphagous than koinobionts (Godfray 1994, Althoff 2003, van Driesche 2004). Pennacchio and Strand (2006) have reviewed developmental strategies in parasitic Hymenoptera. Host suitability can be controlled by a range of other attributes. Grandgirard et al. (2006) determined that indigenous cicadellids of French Polynesia differed sufficiently from the exotic pest Homalodisca vitripennis (Germar) in size, ecology, and oviposition biology to be at no risk from an introduced mymarid egg parasitoid.
Fungal pathogens have been favoured over other plant pathogens for weed biocontrol, partly because they often have the required host-specificity. The most commonly used are biotrophic or obligate species that are generally host specific and debilitate vigorously growing plants by accessing nutrients directly from living tissue (Morin et al. 2006). These include the rusts and smuts and are analogous to koinobiont parasitoids. The recent control of mistflower (Ageratina riparia) using Entyloma ageratinae is an example (Barton et al. 2007). More facultative species that kill cells and then utilise dead plant material have also been used (Morin et al. 2006), and these are analogous to idiobionts. The biological control programme of old man's beard (Clematis vitalba) using Phoma clematidina is an example (Gourlay et al. 2000). Discussion about various aspects of the use of fungi as both classical and augmentative biological control agents for insects can be found in the book edited by Butt et al. (2002).
Host finding cues and behaviour
Successful oviposition by either phytophagous insects or parasitoids follows host-searching behaviour that brings the female to the host. Parasitoids must locate hosts in a complicated and heterogeneous environment and on finding a possible host must make a range of reproductive decisions. The cues are often chemical stimuli emitted by the host insect and/or the host plant. Parasitoids tend to use the odour of invertebrate hosts only for short distance detection. While these direct stimuli are highly specific to the identity of the host, these make up only a small proportion of the biomass within the habitat, and so stimuli are difficult to detect at a distance. Instead, parasitoids often use chemicals associated with the host plant of the target, or the chemicals associated with insect damage to the host plant, insect saliva or insect frass to orient over longer distances. These indirect stimuli are easier to detect, but less reliable as an indicator of the presence of the true host. Turlings et al. (1991) found that these tritrophic stimuli engendered stronger behavioural responses in Cotesia marginiventris than the presence of its host. The response of parasitoids to mixtures of volatiles from plant and host can be complex.
The variety of behavioural cues seems endless. Some parasitoids orient to host pheromones (Godfray 1994), whereas Megarhyssa spp. react to stimuli from the fungus always associated with Sirex larvae, but not to the host larva itself (Spradberry 1970). Physical limitations such as size of nest hole can also influence which hosts are available to a parasitoid (Sands and Van Driesche 2004).
Godfray (1994) divides research into host location by parasitoids into two schools:
- behavioural mechanisms or physiology: this approach has successfully revealed the complexity of the cues used in host location, how these can vary, and the importance of learning;
- behavioural ecology: optimal foraging theory predicts feeding behaviour on the assumption of optimisation by natural selection.
Others (Vet and Dicke 1992) moved on from the concept of a 'hierarchy' of stimuli leading to parasitism (Vinson 1998) and proposed that a newly emerged parasitoid has a range of innate 'response potentials' to different stimuli and at any one time will respond to the stimulus with the greatest response potential. This makes response to hosts variable depending on where the parasitoid is, and sometimes otherwise strong behaviours might be bypassed. The response potentials can vary with experience, so that older parasitoids may respond differently from those that are newly emerged, and may differ depending on where the parasitoid finds itself. Responses vary with previous experience and between individuals and populations, on different plant hosts, and even with presence of other parasitoids. Many examples illustrate the extreme flexibility many parasitoids possess in the incorporation of new stimulus responses into their behavioural repertoire. The frass or cocoon of a Microplitis croceipes entrains the host-finding behaviour of the emerging adult, while other parasitoids are unaffected by their larval environment. This ability of parasitoids to learn the characteristics of their development site in this way is a means by which information can be inherited non-genetically � a form of cultural transmission (Godfray 1994). Once a host is found, parasitoids use a variety of optimal foraging strategies and patch leaving rules (Godfray 1994).
These are just a few of the myriad examples of how parasitoid behaviour can vary not only between but within species. Examples illustrate the extreme flexibility many parasitoids possess in the incorporation of new stimulus responses into their behavioural repertoire. Understanding how the candidate parasitoid behaves in different habitats and in different assemblages is important to the design and interpretation of reliable host range tests. Although difficult to design, oviposition tests should strive to include host-finding as well as host-acceptance behaviour (Barratt 2004, van Driesche and Murray 2004, Babendreier et al. 2005). The design of host range tests for parasitoids is discussed elsewhere.
For some parasitoids oviposition occurs away from the host stage. For example, the planidia and triungulin larvae of parasitic Diptera are either ingested by the foraging larva or are actively carried to the host. These agents are rare in biological control programmes and their behaviour is not discussed here.
It is tempting to think that predatory arthropods have a wider host range and therefore have less complex host-finding mechanisms. However, some coccinellids are relatively host specific, and may well be governed by similarly complex interactions. Causton (2004) noted reports that Icerya purchasi Maskell sequesters different alkaloids from different host plants, and this leads to different developmental performance in its predator Rodolia cardinalis Mulsant.
Habitat specificity, habitat suitability, and agent mobility
Natural enemies do not co-occur universally with their hosts. The distribution of agents is mediated not only by the presence of the host but also by a wide range of physical and biological requirements. The likelihood of non-target effects is therefore dependent on not only the host-specificity of the natural enemy, but also its ability to persist alongside susceptible hosts. Identification of well-documented requirements that limit distribution might allow an applicant to argue that the ecological host range of a proposed control agent was smaller than the fundamental host range. This would occur if the distribution of at risk non-target species did not overlap with the predicted distribution of the control agent, or that agents may perform poorly in that environment. Any claim that a host within the fundamental host range of an agent was excluded from the ecological host range on the grounds of habitat specificity would need to be well-supported.
Many factors other than the presence of a host limit the distribution of parasitoids and herbivores, but the most obvious is climate. Insects are poikilothermic, and absolutely dependent on ambient temperature for their continued development and existence. No biocontrol agent can permanently colonise a habitat outside the lethal temperature range of any of its life stages. Distribution of populations may therefore be limited by elevation or latitude, but predicting distribution accurately is difficult without adequate information about the climate preferences of agents and potential non-target hosts. When Microctonus hyperodae was introduced to New Zealand for the control of the lucerne pest Listronotus bonariensis, it was considered unlikely that the parasitoid would move from the lowland disturbed cropping environment to higher elevation native grasslands. However, parasitoids have colonised such areas and limited attack on non-target weevils has been observed in this cold habitat (Barratt et al. 1997, Barratt 2004).
Distribution can be mediated by rainfall or aridity, crop systems, forest type, aspect, plant species, plant community, and even physical characteristics such as nest hole size (Sands and Van Driesche 2004). Benson et al. (2003) found that Cotesia glomerata foraged in sunny open areas but not in the shade. This is why the brassica crop pest Pieris rapae is attacked in NE U.S.A. but the related forest species P. virginiana is not. Beard and Walter (2001) found that a species of a predatory phytoseid mite was associated with one particular tree species. Kitt and Keller (1998) showed by wind tunnel experiments that only aphids found on rose bushes would be at risk from Aphidius rosae Haliday.
Romeis et al. (2005) reported that parasitism levels by Trichogramma varied greatly among different habitats, plants or plant structures on which the host eggs were located. The mechanisms for this variation included plant spacing, plant structure, plant surface structure and chemistry, plant volatiles and plant colour. In addition, plants can affect parasitoid behaviour and activity by providing carbohydrate food sources such as nectar to the adult wasps, and by affecting the nutritional quality of the host eggs for progeny development. Rutledge and Wiedenmann (1999) tested the responses of three Cotesia species (Braconidae) that parasitise stem-boring Lepidoptera to volatiles from a range of host and non-host grasses and volatiles from non-grass, non-host plants. The three species were differentially attracted to grasses in a pattern consistent with field host records. They suggested that testing preference for habitat plants could help to determine the ecological host range of some parasitoids, assist in matching these to target crops, as well as streamlining testing for impacts against non-target species. Specificity can even be restricted to microsites such as plant structures. Duan and Messing (1998) showed that opiine braconids introduced to Hawaii for fruit fly control only attacked tephritid larvae in fruit or fruit-like structures, and caused no non-target effects on those in flowerhead-gall infesting tephritids. Parasitoids of leafmining species often attack a wide range of species, but the realised host range can be limited by the host plants on which those leafminers feed (Askew 1994).
Habitat specificity does not allow control agent populations to persist. However, susceptible non-target plants may still be at temporary risk if the control agent can disperse out of acceptable habitats in sufficient numbers to cause damage to non-target hosts (spill over). Withers et al. (2008) detected huge populations of the moth Cydia succedana in a valley 4 km from the nearest stand of known hosts. There was no evidence of damage to local non-target plants that might have generated this population. Dispersal monitoring eventually indicated that this large population had accumulated in the valley passively on the prevailing wind. The valley was not a suitable permanent habitat for the moth, but had a susceptible non-target species been present in the valley, significant 'spill over' damage may have resulted as a result of this dispersal.
Even when habitat characteristics allow persistence, population growth and hence any potential risk to non-target plants is affected by environmental conditions. In marginal habitats annual climate patterns might determine oviposition rates and lifetime fecundity, the number of generations in a year, and the onset and cessation of facultative diapause or aestivation. Julien et al. (1995) compared the potential ranges of both alligator weed Alternanthera philoxeroides and its chrysomelid control agent Agasicles hygrophila. The model predicted where the agent would have some impact on the weed and those areas where beetle damage would be little or absent. Where agent and host have different temperature thresholds for development and activity the effect of climate on relative growth rates and reproductive rates may well determine whether non-target impacts will be significant or not.
Native range - experimental confirmation of host range
Applicants to the EPA must demonstrate the range of hosts that a new organism will exploit once released in New Zealand and assess the direct risks of those interactions. It is theoretically possible that the behaviour of the organism in the field in its native range could provide sufficient information to do this, but it is more likely that experimental evaluation of the organism's host range testing will be required to achieve this. The purpose of conducting host range tests on a potential biological control agent is to avoid introducing an agent that might cause significant economic or environmental damage while avoiding the unnecessary rejection of a potentially effective agent. Host-specificity testing remains the most important method for predicting the safety of both weed (Briese 2006) and arthropod (van Lenteren et al. 2006, van Lenteren et al. 2006) control agents.
Host-range studies to predict possible non-target effects of biological control agents for weeds were first conducted in the 1920's (McFadyen 1998). Based largely on a retrospective study of almost 50 years of such research, Wapshere (1974) proposed a strategy for evaluating the safety of organisms for biological weed control in which plants of decreasing relatedness are presented to the control agent until no further attack is observed. The pattern of attack revealed by this process defines the taxonomic limit (species, genus, etc) within which the host range lies. This hypothesis has become standard practice throughout the world for determining the fundamental host range of potential control agents for weeds. Wapshere's original paradigm has proven to be robust, and appears to be equally useful in predicting the host range of biotrophic pathogens (Morin and Edwards 2006).
There have been few examples recorded where these standardised procedures for estimating field host range of arthropod or fungal control agents have failed (Fowler et al. 2000, Pemberton 2000, Barton 2004). In New Zealand, a recent survey of the field host range of most of the agents introduced for the biological control of weeds indicated no cases where the recorded host-range was not, or could not have been predicted by appropriate application of centrifugal phylogenetic testing before the agents were released (Paynter et al. 2004).
Historically, testing of control agents for arthropod pest control has been restricted to checking that the parasitoid or predator attacks the target host, is not a hyperparasitoid of other natural enemy of the host, and does not attack biocontrol agents of other pests. van Driesche and Hoddle (2000) reviewed the techniques available at that time and recommended that the non-target species tested should include not only insects that are closely-related to the host, but also arthropods of special ecological or scientific interest even if distantly related, including biological control agents.
The process of determining the host range of a parasitoid experimentally is complicated by other issues, and sorting out the framework for predicting host range is a big theoretical issue (Bigler et al. 2006). Host range testing of parasitoids is conducted despite these constraints. van Driesche and Hoddle (2000) and Kuhlmann et al. (2005) list and discuss a range of programmes conducted over the last 24 years that have assessed the host range of parasitoids prior to release. While wider host-testing of parasitoids developed for arthropod control has been carried out in New Zealand since the 1970's (Hill et al. 1985), this has not been standard practice in other parts of the world. Recent New Zealand exemplars include Cameron and Walker (1989), Cameron et al. (1995), Stufkens et al. (1994), Cameron and Walker (1997), Barratt et al. (1997), Froud and Stevens (1998), ERMA New Zealand (2000), Charles (2001) and Berndt et al. (2007). Some recent testing programmes for control agents for weed control in New Zealand are described by Hill et al. (2001), Hill et al. (2001), Hill and Gourlay (2002) and Grosskopf et al. (2002).
The factors influencing host selection in natural enemies have already been reviewed in this section. The next two sections review how species are selected for testing, and the experimental techniques used to evaluate these influences on agent safety.
Selecting test species
Every biological control project is unique, and in practice there are no universal rules for selecting which plant or arthropod species should be tested to evaluate the safety of a control agent, or how potential impacts on ecosystems should be assessed. Even without such a fixed framework, applicants to the EPA are required to present a coherent and convincing case that the proposed control agent will not displace or significantly impact on valued flora and fauna. In the past ten years ERMA New Zealand accepted evidence based on as few as two test species (Froud and Stevens 1998) and as many as 46 (see decision on application to ERMA New Zealand NOR02001, go to Documents and Decision [http://www.epa.govt.nz/search-databases/Pages/applications-details.aspx?appID=NOR02001#]) or more. It is up to the applicant to design the risk assessment on knowledge of the biology of the target and its natural enemy, and on ecological and biological control theory.
The strategy proposed by Wapshere (1974) to define the host range of a natural enemy is based on the principle that a plant more closely related to the target weed will present more feeding cues and fewer feeding deterrents to a specialist arthropod herbivore than a plant that is less closely related to the target plants, and will therefore be more likely to be a host for that insect. He formalised this into a procedure called 'centrifugal-phylogenetic method, and this has been the basis for the selection of plants to test ever since. To this he added a set of 'safeguard' criteria for assessing risk to plants that would not be selected on phylogenetic grounds alone, but for which host relationships were not well known. These safeguards were:
- cultivated plants botanically related to the target weed;
- cultivated plants for which there is little or no entomological or mycological knowledge;
- cultivated plants which for evolutionary, geographic or climatic reasons have not been extensively exposed to the candidate agent;
- cultivated plants known to be attacked by organisms closely related to the candidate agent;
- any plants on which the candidate agent has been previously recorded.
Perceptions of risk have changed in the 30 years since Wapshere proposed his protocol. There is greater awareness of the risk posed by colonisation of non-target plants in the receiving range, the possibility of control agents displaying taxonomically 'disjunct' host ranges (Kay and Hill in press), and unacceptable temporary damage caused by 'spill over' feeding (increased awareness of the indirect risks posed by control agents are discussed below).
Briese (2003) suggested three improvements to the criteria for test plant selection that would contribute to better risk analysis:
- use true phylogenetic distance (rather than taxonomic distance) to measure relatedness between plant species;
- consider the degree of biogeographic overlap (this is less relevant for a small landmass like New Zealand, but presumably the same argument can be made for ecoclimatic overlap);
- consider the degree of ecological similarity between potential non-targets.
Despite these recommendations for improvement, it is generally acknowledged that the selection of test plant species to predict the likely host range of weed control agents is well supported by theory.
For oligophagous parasitoids, choosing which species to test, and which factors to incorporate into test designs is more problematic. As it is for weed control agents, the selection of species on the basis of phylogenetic similarity is generally considered a good starting point (van Driesche and Hoddle 1997, Kuhlmann et al. 2005). However, a number of factors make this approach less reliable for parasitoids and predators than it is for weed control agents, including:
- the phylogeny of host arthropods is less well known than the phylogeny of
host plants (
); - most plants are known, but many potential non-target arthropod species are as yet unknown. In New Zealand these may form the bulk of potential species to test (Barratt et al. 2000);
- although known from collection or literature records, some species that should be tested may be unobtainable or impossible to rear (Barratt 2004);
- the host-range of even closely related parasitoids can be disjunct. For many taxonomic groups this pattern is not sufficiently systematic to support an hypothesis as robust as the Wapshere protocols;
- host specificity that appears to be related to taxonomy may instead be related to a correlate of taxonomy, such as the tendency of related species to share habitats (van Driesche and Hoddle 1997);
- many parasitoid groups have predictable host associations but others do not (Sands and Van Driesche 2004). Even amongst predatory arthropods the host range of one species does not reliably mirror the host range of a congener (Causton 2004);
- there are more arthropods from which to choose test plants than there are plants, and so tests of potential hosts of parasitoids are more difficult and expensive than hosts of phytophages;
- it is easier to grow and manipulate plants than insect cultures, and experiments involving insects are more difficult logistically. A list of insects to test must therefore be short;
- rather than being driven largely by phylogenetic similarity, the factors that determine host selection of parasitoids and predators are varied and layered, and difficult to isolate in simple tests (
).
Kuhlmann et al. (2006) described and analysed a range of host-range testing programmes and listed five criteria that have been used to select species for testing against invertebrate biological control agents:
- ecological similarity between host and test species � overlapping geographic range, feeding niche, habitat preference;
- phylogenetic affinity between host and test species - predicted by taxonomic relatedness;
- socio-economic importance of the possible test species� conservation importance, commercial importance, role as a beneficial;
- biological similarity between host and test species � known host range, phenological overlap with the target, dispersal capability, morphological similarity, behavioural similarity, overlap of physiological host range of other agents, response to host plant (Kitt and Keller 1998), similarity of host plant structure (Duan and Messing 1998);
- availability of test species for testing, and the potential for using surrogates.
The information supporting these decisions usually came from the literature, and rarely from first hand experience in the home range of the agent. Babendreier et al. (2005) and Babendreier et al. (2006) also review the criteria for selecting test species. They noted that in many projects only congeners are tested, and recommended improving the margin for error by choosing a wider array of test species.
Recently a decision support system designed to aid the selection of non-target species (NTS) for testing with proposed biological control agents was developed here in New Zealand. The PRONTI (priority ranking of non-target invertebrates) tool was developed for risk assessment biosafety testing of transgenic plants (Todd et al. 2008, Malone et al. 2010, Barratt et al. 2013), and has been adapted and tested for use with entomophagous biological control agents (Todd et al. 2015, Barratt et al. 2016). This was possible because the criteria used to select NTS for biosafety testing with these organisms are fundamentally the same (i.e., the risk posed by the new organism, and the value and testability of the NTS).
Essentially, PRONTI uses a mathematical model and a database of information on NTS, called the Eco Invertebase, to prioritise species for biosafety testing with each proposed biological control agent (Todd et al. 2015). The Eco Invertebase holds information (i.e., taxonomy, food web, biomass, ecology, anthropocentric value and testability data) on more than 1500 invertebrate taxa found in New Zealand. The PRONTI model prioritises the taxa in the Eco Invertebase by applying five selection criteria: (1) the potential hazard posed by the biological control agent to each NTS; (2) the potential degree of exposure of each NTS to the agent; (3) the hypothetical ecological impact that may result from the exposure of the NTS to the hazard posed by the agent; (4) the estimated economic, social and cultural value of each NTS; and (5) the assessed ability to conduct tests with the NTS.
The model output provides information on how each taxon compares with all other taxa in the list using the five criteria. It also provides a measure of certainty of each taxon�s ranking. The model was partially validated by comparing the PRONTI list for the predatory wasp Polistes chinensis with the prey of this species in the field (Todd et al. 2015; see figure below).
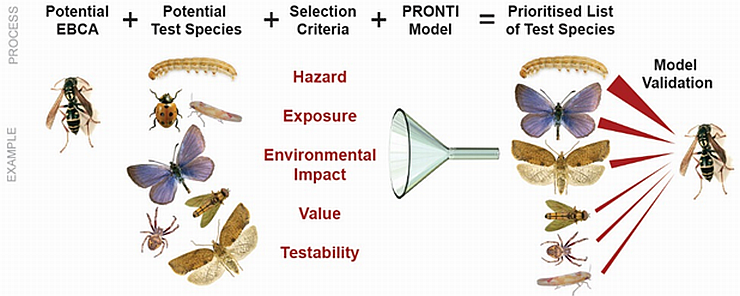
The PRONTI process. The example illustrates the process using the hypothetical biological control agent, the predatory wasp Polistes chinensis, and partial validation of the model through a comparison of the high risk species with the prey items collected from P. chinensis colonies in the field.
© Copyright Plant & Food Research, used with permission.
Currently, PRONTI is undergoing a �live test� with an agent which is proposed for release in New Zealand: Eadya paropsidis Huddleston & Short (Hymenoptera: Braconidae) to target the forestry pest Paropsis charybdis St�l (Coleoptera: Chrysomelidae). Again, PRONTI will be used to rank at least 130 NTS, and the resulting list will be compared with that produced using the currently accepted methodology (Withers et al. 2015). If the PRONTI list is considered to be informative for New Zealand�s Environmental Protection Authority and other decision makers, then PRONTI may become recommended best practice for New Zealand�s biocontrol agent researchers in future. In the meantime if you are interested in testing this tool for your project, contact [email protected]
Testing methods
The styles of tests that can be employed are well reviewed elsewhere in BIREA. van Driesche and Reardon (2004) edited a guide to best practice for assessing the host range of predators and parasitoids. That publication is not reviewed here. Heard and Van Klinken (1998), Sheppard (1999) and Spafford-Jacob and Briese (2003) have reviewed the use of different designs for host range testing of phytophages for weed control. van Driesche and Murray (2004) have reviewed the strengths and weaknesses of the designs that can be employed in host range testing parasitoids and reached the general conclusion that as far as possible the test regime adopted should model the ecological context in which the agents will interact with potential hosts. This view is also implicit in the selection criteria suggested by Kuhlmann et al. (2006).
In the view of Babendreier et al. (2006), the purpose of host range testing is to determine the ecological host range of the natural enemy. This seems ambitious, and they acknowledge that designing tests for this purpose is difficult. An alternative view is that host range testing provides an opportunity to define the physiological or fundamental host range, and by the use of a range of test designs, to provide additional information with which to predict the likely field host range. This approach seems more likely to be successful. A test design array should take into account the following principles:
- Factors within test designs, such as the size of the arena or the presence of plant material, may either reduce or increase the acceptability of hosts to natural enemies (Babendreier et al. 2005). At least one test (usually a simple, well controlled 'no choice' test in conditions that are highly suitable for the agent) must create a 'maximum challenge' to the susceptibility of the host to create a reliable physiological host range (Hill 1999, Withers and Barton-Browne 2004). However, it is important for reviewers to distinguish the physiological host range revealed by such tests from the likely field host range.
- 'Choice' tests are often used because this design represents field conditions more realistically than 'no choice' tests. The value of 'choice' tests is debated (Barratt et al. 2006), and depending on factors such as agent size and mobility, 'no choice' may well be the more realistic design (Hill 1999).
- Natural enemies can respond differently to their host depending on age and physiological state. A newly emerged natural enemy may have a different intensity of response than an aged individual. For example, Kitt and Keller (1998) found that older Aphidius rosae adults were more selective than newly emerged individuals. A satiated agent may attack fewer hosts than a deprived agent. The duration of a test may therefore influence its outcome. Test designs must consider these possibilities. The issues posed by time and behaviour-dependent effects are reviewed by Withers and Barton-Browne (2004).
- Marohasy (1998) reviewed the design and interpretation of host-specificity tests for weed biological control agents and recommended that tests should be designed to take more account of agent behaviour. She suggested methods for measuring the rank order of acceptability of test species, and the time taken before an agent will accept a lower-ranked plant species as additional measures of host acceptance. These novel approaches may well have application in the testing of arthropod control agents.
- Hosts must be presented in tests in the correct or most acceptable stage, and it is important that the target host is represented in each design (Barratt et al. 1999, Babendreier et al. 2005).
- In their native range the host range of parasitoids and phytophages can vary between populations (see discussion about biotypic and geographic variation above). Best practice in New Zealand now dictates that only those populations that have been tested should be released. Later releases of populations from other sources should be subject to confirmation of that host range by limited testing (Withers et al. 2008).
van Lenteren et al. (2006) have summarised the issues that should be considered when designing host specificity tests on arthropod natural enemies, and propose a methodology for addressing those issues. Their methodology is very clear and well-annotated, and provides a sound baseline for the development of a testing regime. However, they acknowledge that a theoretical and methodological background for such a testing regime for arthropod natural enemies is not yet mature. They summarise what information should be brought to bear on a test design:
- know the foraging behaviour of the natural enemy;
- know the influence of quality and rearing conditions of the host plant, host and natural enemy. These should be described in detail to trace the effects of conditioning, learning and multitrophic communication on results;
- understand the effect of diapause state of both host and parasitoid;
- avoid hosts with infections;
- recognise behavioural variation in natural enemies and identify the source (genetic, phenotypic, physiological);
- understand the genetic characteristics of the parasitoid and match strains to the host;
- consider the environment of the test and how this might affect associative learning leading to changes in relative behaviours of host and parasitoid;
- be aware of the influence of biotypes and multitrophic stimuli on design (van Lenteren et al. 2006, van Lenteren et al. 2006).
Berndt et al. (2007) provide an example of how to develop a test design.
What we do know
The introduction of exotic biological control agents is not an intrinsically safe operation, but one that is made safe by the way it is practiced (McClay and Balciunas 2005). This is understood by every responsible biological control practitioner, and is reflected in the wording of the HSNO Act, and in the Methodology Order that interprets it. One of the guiding principles of the Act is the safeguarding of the life-supporting capacity of ecosystems, and in their deliberations the EPA must take account of the inherent value of ecosystems. The Act requires the EPA to reject an application if it would result in significant deterioration of a natural habitat or significant displacement of a native species in its natural habitat. Applicants must supply sufficient information for the EPA to be able to make these judgements.
A new organism will always modify the ecosystem into which it is introduced, however slightly. It is therefore wrong to suggest that biological control has no effect on communities and ecosystem processes. Hoddle (2004) concluded that adding exotic organisms to an ecosystem has three effects: the links in food webs that connect populations are permanently changed; pest density population dynamics are altered; and changes in the relative dynamics of populations cause changes in community structure. van Lenteren et al. (2006) review the range of potential non-target effects that might be modified by the introduction of an exotic biological control agent. The control agent may attack a species other than the target pest and affect its abundance. The possible consequences of direct non-target attack have already been reviewed. Indirect non-target effects occur when the presence of the agent changes community structure by means other than consumption. Resource competition occurs when two organisms exploit and divide a single resource. This may reduce the population of the displaced species, sometimes to local extinction, but it has rarely if ever led to species extinction (van Lenteren et al. 2006). Parasitoids and predators may be able to attack other natural enemies of the target; this is termed intraguild predation or facultative hyperparasitism (van Veen et al. 2006). The natural enemy may itself be food for resident natural enemies. This is especially so for biological control agents for weeds. This intrusion into the ecosystem is called enrichment, and may provide a food subsidy for parasitoids and predators within the ecosystem. If this leads to increased natural enemy populations, and increased mortality of another resident host species, then the introduction has caused apparent competition between the introduced and the resident hosts. These indirect effects can be either via a host (bottom-up, McEvoy and Coombs 1999), or top-down via a higher trophic level (Holt and Hochberg 2001). The density dependent regulation of ineffectual generalists does not occur under apparent competition and non-target populations can decline (Hoddle 2004).
Messing et al. (2006) reviewed the literature on indirect non-target effects, and attempted to synthesise the styles of interaction described. Pearson and Callaway (2005) have also reviewed the literature on indirect non-target effects.
What we don't know
Much of the recent research into non-target effects has concentrated on two-species interactions compared with the more subtle indirect interactions such as competition and apparent competition (Messing et al. 2006). In part this is because despite mounting information about community dynamics and how to study them, the inherent complexity and uncertainty of interactions within receiving communities makes rational decision-making about non-target effects difficult (van Lenteren et al. 2006). Babendreier et al. (2006) have reviewed the various international attempts at developing regulations for biological control, and detail the information requirements of the OECD on environmental risk assessment of biological control agents. Although information on competition and other indirect effects are now required by regulators like the OECD, it unclear how this will be achieved (Babendreier et al. 2006. With relatively few exceptions, studies reporting non-target effects provide information about effects at the level of the individual, such as percentage damage and percentage mortality. Messing et al. (2006) and others propose that evaluation of indirect effects should concentrate on population and community level effects rather than effects on individuals, and need to be field-based and long-term, to isolate temporal and spatial variation. Lockwood (2000) also considered that the conservation of 'species' as a basis for non-target impact assessment lacked depth. He made the case that a population is not a collection of individuals but the energy and nutrient flows that manifest through the individual. For this reason, in assessing impacts the emphasis should be on population and community processes, such as reproduction, competition, consumption, not on population size. Babendreier et al. (2005) suggested that we need to develop better a priori definitions of what a significant non-target effect is, and we should aspire to baseline studies before release. Such aspirations are difficult in the research-funding environment that we work in New Zealand, but this important suggestion should at least focus applicant's minds on potential issues.
How important are these indirect non-target impacts to ecosystem function? The number of deliberate introductions of exotic organisms for the purpose of biological control is small compared with the powerful influence of invasions caused by the accidental introduction of exotic organisms or for the deliberate introduction for other purposes (van Driesche and Hoddle 1997). In terms of relative risk, the role of modern biological control of pests and weeds in environmental change in New Zealand is therefore likely to be small. In a large survey of database information Lynch and Thomas (2000) found that only 1.5% of the 5000 projects they reviewed recorded adverse effects, but they acknowledged that this information was not well represented in the data. Of that 1.5%, only 10% appeared to have effects at the population level. For weed control projects in New Zealand the adverse effects are likely to be less frequent than this (Fowler et al. 2003).
There is debate about how prevalent significant adverse non-target effects are, but there is no debate that these sometimes occur. Louda et al. (2003) reviewed the known examples of adverse effects and provide views on how cases like these might be avoided in future. Simberloff and Stiling (1996) recommended care in the use of biological control because even if we could predict events such as extinction of populations, it is not obvious how to assign value. Beyond the fact that we cannot predict ripples in communities and ecosystems (let alone species) how much is the very existence of these entities worth? The uncertainty is made more acute by the irreversible nature of biological control, and ability of agents to spread. They argued that while biological control can be justified by the high threat of invasives in native communities this does not abrogate the responsibility of biological control practitioners to take care, a recommendation that seems entirely fair.
Hoddle (2004) stated that the published literature suggests that direct attacks on non-target organisms by introduced control agents can be minimised by selecting agents that are both host and habitat specific. Hawkins et al. (1999, cited in Hoddle 2004) suggest that the trophic relationships of successful biological control projects are less reticulate than those in food webs of natural systems. Less successful projects have more impact on food webs because the agent becomes permanently abundant. The best protection against subtle non-target effects is to maximise the host specificity and efficacy of agents (van Lenteren et al. 2006 and others).
Biological control practitioners who wish to introduce a control agent to New Zealand are required to assess the potential for indirect non-target impacts when it is clear that a useful framework is only just in existence. Babendreier et al. (2006) point out that the OECD framework now requires information on the indirect effects of prospective control agents, but how this can be achieved is unclear.
Messing et al. (2006) concluded that practitioners and regulators have a daunting task, as frameworks to guide prediction are rudimentary, and yet concrete and responsible decisions need to be made. They also stressed that it is essential that regulators do not extrapolate from observations of feeding or parasitism in lab or field to population effects. The EPA must operate within the limitations of our understanding of community dynamics in New Zealand ecosystems. In the face of this complexity, applicants will be asked to provide all available information, common sense interpretation of that information, and an appropriate risk assessment based on that interpretation. The following tools have been used in the interpretation of community interactions, and the examples may assist in future risk assessments.
Food webs in native and exotic ranges
Food webs provide a description of the components of a community and of the interactions between those components. Binary food webs use the presence or absence of interactions to illustrate linkages between species in a community. Simple food webs might be easily constructed in simple ecosystems such as crops, but in not in more complex environments. In natural habitats for example, not only are the interactions highly complex, but many of the components may be unknown to science. Construction of simple food webs may therefore require considerable research. Fully quantitative food webs provide a measure of the relative frequency of interactions, as well as the relative abundance of the components. All quantitative food webs are data hungry and expensive to prepare. The value and limitations of food webs in describing the processes that structure communities are reviewed by van Veen et al. (2006). Food webs constructed in the native range could help predict the host range and parasitoid susceptibility of the proposed agents. Food webs constructed in the receiving range could help identify potential parasitoids and competitors for the proposed agent.
Willis and Memmott (2005) demonstrated how food webs can be used to show the interactions between plants, seed-feeding insects and their parasitoids, and to generate testable hypotheses regarding indirect interactions between introduced agents and non-target species. Their model was the weed, bitou bush, Chrysanthemoides monilifera ssp. rotundata, and a biocontrol agent for this weed in Australia, the tephritid fly, Mesoclanis polana. The food webs revealed how a highly host-specific biocontrol agent such as M. polana has the potential to change community structure by increasing the abundance of native parasitoids. The webs also suggest that indirect interactions between M. polana and native non-target species were possible, mediated by shared parasitoids.
Hopper and Wajnberg (2006) described the food web based on the Lepidoptera of Alakai Swamp on Kauai, Hawaii. They showed that 83% of parasitoids reared from native moths were biological control agents (all introduced prior to 1945), 14% were accidental immigrants, and 3% were native species. Parasitism by biological control agents reached 28% in some species of moth. However, the authors could not assert what this meant for the population dynamics of the species attacked, and hence for the community dynamics in the swamp, or whether species had already been lost from the community.
Munro and Henderson (2002) used quantitative webs to interpret the complex interactions between native parasitoids of native Lepidoptera and the introduced species Trigonospila brevifacies (Hardy) in New Zealand. Its host range overlaps with 12 native and one introduced parasitoid species, and it parasitised more species of Tortricidae than other parasitoids at the North Island forest sites surveyed. All native parasitoid species were less abundant than T. brevifacies and this species comprised 15.6 - 79.5% of the parasitoid load per species.
Food webs summarise the trophic interactions in a community plus give insights into indirect but trophically-mediated interactions such as resource exploitation and apparent competition. In food webs based on phytophagous insects there is huge scope for trait mediated indirect effects (van Veen et al. 2006). There is increasing awareness that other interactions may be equally important, particularly where behaviour of one species is modified by another. For example, the impact of kairomones on parasitoid behaviour is not captured in food webs. Infochemical interaction webs might be a natural counterpart to trophic webs (Vet and Godfray 2008).
Food webs have value for visualising interactions in communities, but as yet do not have sufficient power to predict how natural enemy interactions structure communities (Hopper and Wajnberg 2006, van Veen et al. 2006).
Lockwood (2000) considered that the conservation of 'species' as a basis for non-target impact assessment lacked depth. He made the case that a population is not a collection of individuals but the energy and nutrient flows that manifest through the individual. For this reason, in assessing impacts the emphasis should be on population and community processes, such processes as reproduction, competition, consumption, not on population size.
Manipulation and surrogate systems
The ability of control agents to attack non-target species has been estimated by exposing susceptible life stages in the field. Babendreier et al. (2006) review a number of such studies.
Retrospective studies
Post-release studies tell us nothing about the risk profile of any particular natural enemy proposed for introduction, and are therefore not strictly relevant to the preparation of any application to the EPA to introduce a new organism. However, such studies are essential if we are to later verify the predictions made in applications, and so build confidence in methodologies for predicting non-target effects. Barratt et al. (2006) review the role of post-release studies in improving pre-release evaluation of the risks of biological control. They acknowledge that no accepted standards yet exist for doing this.
A high degree of host specificity limits the number of interactions that a control agent will have in its new environment (
. The reliability of host- range testing is therefore an important issue in minimising non-target community effects. Comparisons of the field host range of natural enemies following release with the predictions of host range testing are becoming more common, and for the most part appear to be validating the predictive capacity of host-range testing. Paynter et al. (2004) described a large survey of the non-target plants of 20 control agents introduced to New Zealand for the biological control of weeds since 1926. Eighteen were either host-specific or the field host range was originally predicted by testing. For two species, Bruchidius villosus (F.) and Cydia succedana (D&S) minor attack on several exotic non-target hosts related to the true host was not predicted by testing regimes. However, the levels of attack are likely to be insignificant to community structure. These results complemented those of Fowler et al. (2003) who found that (with the exceptions mentioned) host range testing for weed control agents in New Zealand has reliably predicted post-release field host range. Hill et al. (2001) predicted that Phytomyza vitalbae, a leaf mining control agent of Clematis vitalba, was likely to occasionally attack leaves of native Clematis species. Paynter et al. (2006) surveyed damage to the leaves of Clematis species throughout New Zealand and found that this prediction was largely correct. There are other examples of post-release field studies that confirm the host range predicted by testing, including Haye et al. (2005) and Coombs (2003). Barratt et al. (2006) reviewed their extensive research into the performance of the weevil parasitoid Microctonus aethiopoides in New Zealand following its release for the control of Sitona discoideus. They have examined how spatial distribution, phenology and habitat affect the propensity of the parasitoid to utilise non-target native weevils, and the consequences for weevil populations.
Barron et al. (2003) used a life table analysis to investigate the effect of the pupal parasitoid Pteromalus puparum (L.) on the population dynamics of the red admiral, Bassaris gonerilla (F.) in New Zealand, over fifty years after the parasitoid was introduced for white butterfly control. They found that although common and apparently damaging, parasitism by P. puparum was less important to the population dynamics of the host than egg parasitism by Telenomus sp. and mortality caused by a second pupal parasitoid, Echthromorpha intricatoria (F.). Such studies could help pinpoint key factors in the population dynamics of important non-target species, and avoid increasing mortality in critical life stages.
Barlow (1999) found no existing population models aimed at predicting non-target impacts but predicted that the demand for such models would grow. However, he pointed out that there is likely to be far less information about non-target species on which to base such models. Barlow et al. (2004) then went on to model the population dynamics of a non-target host in the presence and absence of the introduced braconid M. aethiopoides. The difference in population impact between habitats was explained by the difference in the intrinsic rate of increase of the non-target species in those habitats.
Non-target impacts can occur only if natural enemies and potential hosts share habitats, e.g. Barratt et al. (2006). Pre-release climate modelling based on the biological characteristics of the agent has the potential to predict such overlaps (see above). Louda et al. (2005) found that the environment influences and can alter host use and population growth, leading to higher than expected direct impacts on the less preferred native host species at several spatial scales. They concluded intensive retrospective ecological studies provide guidance for future assessment of candidate biological control agent dynamics and impacts.
References
Althoff D.M. (2003). Does parasitoid attack strategy influence host specificity? A test with New World braconids. Ecological Entomology 28: 500-502.
Askew R.R. (1994). Parasitoids of leaf-mining Lepidoptera: what determines their host ranges? Pp. 177-202 In: Parasitoid community ecology, B.A. Hawkins and W. Sheehan (Ed.) Oxford University Press, Oxford
Babendreier D., Bigler F. and Kuhlmann U. (2005). Methods Used to Assess Non-target Effects of Invertebrate Biological Control Agents of Arthropod Pests. BioControl 50: 821-870.
Babendreier D., Bigler F. and Kuhlmann U. (2006). Current status and constraints in the assessment of non-target effects. Pp. 1-13 In: Environmental impact of invertebrates for biological control of arthropods � methods and risk assessment, F. , F. Bigler, D. Babendreier and U. Kuhlmann (Ed.) CABI Publishing, Wallingford, UK
Barlow N.D. (1999). Models in biological control: a field guide Pp. 43-70 In: Theoretical approaches to biological control, B.A. Hawkins and H.V. Cornell (Ed.) Cambridge University Press UK.
Barlow N.D. and Goldson S.L. (1993). A modelling analysis of the successful biological control of Sitona discoideus (Coleoptera: Curculionidae) by Microctonus aethiopoides (Hymenoptera: Braconidae) in New Zealand. Journal of Applied Ecology 30: 165-179
Barlow N.D., Barratt B.I.P., Ferguson C.M. and Barron M.C. (2004). Using models to estimate parasitoid impacts on non-target host abundance. Environmental Entomology 33: 941-948.
Barlow N.D., Goldson S.L. and McNeill M.R. (1994). A prospective model for the phenology of Microctonus hyperodae (Hymenoptera: Braconidae), a potential biological control agent of Argentine stem weevil in New Zealand. Biocontrol Science and Technology 4: 375-386.
Barlow N.D., Kean J.M. and Goldson S.L. (2002). Biological control lessons from modeling of New Zealand successes and failures. Proceedings of the First International Symposium on Biological Control of Arthropods: 105-107.
Barratt B.I.P. (2004). Microctonus parasitoids and New Zealand weevils: comparing laboratory estimates of host ranges to realized host ranges. Pp. 103-120 In: Assessing host ranges for parasitoids and predators used for classical biological control: A guide to best practice, R.G. Van Driesche and R. Reardon (Ed.) USDA Forest Service, Morgantown, West Virginia.
Barratt B.I.P., Blossey B. and Hokkanen H.M.T. (2006). Post-release evaluation of non-target effects of biological control agents. Pp. 166-186 In: Environmental Impact of Arthropod Biological Control: Methods and Risk Assessment, U. Kuhlmann, F. Bigler and D. Babendreier (Ed.) CABI Bioscience, Delemont, Switzerland.
Barratt B.I.P., Evans A.A., Ferguson C.M., Barker G.M., McNeill M.R. and Phillips C.B. (1997). Laboratory nontarget host range of the introduced parasitoids Microctonus aethiopoides and Microctonus hyperodae (Hymenoptera: Braconidae) compared with field parasitism in New Zealand. Environmental Entomology 26: 694-702.
Barratt B.I.P., Evans A.A., Ferguson C.M., McNeill M.R. and Addison P. (2000). Phenology of native weevils (Coleoptera: Curculionidae) in New Zealand pastures and parasitism by the introduced braconid, Microctonus aethiopoides Loan (Hymenoptera: Braconidae). New Zealand Journal of Zoology 27: 93-110.
Barratt B.I.P., Ferguson C.M., Bixley A.S., Crook K.E., Barton D.M. and Johnstone P.D. (2007). Field parasitism of nontarget weevil species (Coleoptera : Curculionidae) by the introduced biological control agent Microctonus aethiopoides Loan (Hymenoptera : Braconidae) over an altitude gradient. Environmental Entomology 36: 826-839
Barratt B.I.P., Ferguson C.M., McNeill M.R. and Goldson S.L. (1999). Parasitoid host specificity testing to predict host range. Pp. 70-83 In: Host specificity testing in Australasia: towards improved assays for biological control, T.M. Withers, L. Barton-Browne and J.N. Stanley (Ed.) CRC for Tropical Pest Management, Brisbane, Australia.
Barratt B.I.P., Howarth F., Withers T.M., Kean J.M. and Ridley G. (2010). Progress in risk assessment for classical biological control. Biological Control 52: (3) 245-254.
Barratt B.I.P., Oberprieler R.G., Ferguson C.M. and Hardwick S. (2005). Parasitism of the lucerne pest Sitona discoideus Gyllenhal (Coleoptera: Curculionidae) and non-target weevils by Microctonus aethiopoides Loan (Hymenoptera: Braconidae) in south-eastern Australia, with an assessment of the taxonomic affinities of non-target hosts of M. aethiopoides recorded from Australia and New Zealand. Australian Journal of Entomology 44: 192-200.
Barratt B.I.P., Todd J. and Malone L.A. (2016). Selecting non-target species for arthropod biological control agent host range testing: evaluation of a novel method. Biological Control 93: 84-92.
Barratt B.I.P., Todd J.H., Ferguson C.M., Crook K., Burgess E.P.J., Barraclough E.I. and Malone L.A. (2013). Biosafety testing of genetically modified ryegrass (Lolium perenne L.) plants using a model for the optimum selection of test invertebrates. Environmental Entomology 42: 820-830
Barron M.C., Barlow N.D. and Wratten S.D. (2003). Non-target parasitism of the endemic New Zealand red admiral butterfly (Bassaris gonerilla) by the introduced biological control agent Pteromalus puparum. Biological Control 27: 329-335.
Barton J.E. (2004). How good are we at predicting the field host-range of fungal pathogens used for classical biological control of weeds? Biological control in agricultural IPM systems 31: 99-122
Barton J.E., Fowler S.V., Gianotti A.F., Winks C.J., de Beurs M., Arnold G.C. and Forrester G. (2007). Successful biological control of mist flower (Ageratina riparia) in New Zealand: Agent establishment, impact and benefits to the native flora. Biological Control 40: 370-385
Beard J.J. and Walter G.H. (2001). Host plant specificity in several species of generalist mite predators. Ecological Entomology 26: 562-570
Benson J., Pasquale A., Van Driesche R. and Elkinton J.S. (2003). Assessment of risk posed by introduced braconid wasps to Pieris virginiensis, a native woodland butterfly in New England. Biological Control 26: 83-93
Berndt L.A., Mansfield S. and Withers T.M. (2007). A method for host range testing of a biological control agent for Uraba lugens. New Zealand Plant Protection 60: 286-290.
Bigler F., Babendreier D. and Kuhlmann U. (2006). Environmental impact of arthropod biological control: methods and risk assessment. Pp. 288. CABI Publishing, Delemont, Switzerland.
Briese D.T. (2003). The centrifugal phylogenetic method used to select plants for host-specificity testing of weed biological control agents: Can and should it be modernised? In: Improving the selection, testing and evaluation of weed biocontrol agents, H. S. Jacob and D. T. Briese (Ed.) CRC for Australian Weed Management Technical Series no. 7, Adelaide, Australia
Briese D.T. (2006). Can an a priori strategy be developed for biological control? The case for Onopordum spp. thistles in Australia. Australian Journal of Entomology 45: 317-323
Buckley Y.M., Rees M. and Paynter Q. (2004). Modelling integrated weed management of an invasive shrub in tropical Australia. Journal of Applied Ecology 41: 547-560
Butt T.M., Jackson C. and Magan N. (2002). Fungi as Biological Control Agents: Progress, Problems and Potential. CABI Publishing, Wallingford, U.K. 390 pp.
Cameron P.J. and Walker G.P. (1989). Status of introduced larval parasitoids of tomato fruitworm. Proceedings of the New Zealand Plant Protection Conference 42: 229-232
Cameron P.J. and Walker G.P. (1997). Host specificity of Cotesia rubecula and Cotesia plutellae, parasitoids of white butterfly and diamondback moth. Pp. 236-241 In: Proceedings of the 50th New Zealand Plant Protection Conference, M. O'Callaghan (Ed.) NZ Plant Protection Society Inc.
Cameron P.J., Hill R.L., Bain J. and Thomas W.P. (1989). A review of biological control of invertebrate pests and weeds in New Zealand 1874-1987. CAB International Wallingford, UK and DSIR, New Zealand.
Cameron P.J., Walker G.P. and Keller M.A. (1995). Introduction of Cotesia rubecula, a parasitoid of white butterfly. Proceedings of the New Zealand Plant Protection Conference 48: 345-347
Causton C.E. (2004). Predicting the field host range of an introduced predator, Rodolia cardinalis Mulsant, in the Galapagos. Pp. 195-239 In: Assessing host ranges for parasitoids and predators used for classical biological control: A guide to best practice, R. G. Van Driesche and R. Reardon (Ed.) USDA Forest Service, Morgantown, West Virginia
Charles J.G. (1998). The settlement of fruit crop arthropod pests and their natural enemies in New Zealand: an historical guide to the future. Biocontrol News and Information 19: 47-58
Charles J.G. (2001). Introduction of a parasitoid for mealybug biocontrol: a case study under new environmental legislation. New Zealand Plant Protection 54: 37-41
Charles J.G. and Allan D.J. (2002). An ecological perspective to host-specificity testing of biocontrol agents. New Zealand Plant Protection 55: 37-41
Coombs M. (2003). Post-release evaluation of Trichopoda giacomellii (Diptera: Tachinidae) for efficacy and non-target effects. Pp. 399-406 In: Proceedings of the 1st International Symposium on Biological Control of Arthropods, Honolulu, Hawaii, 14-18 January 2002, R. G. Van Driesche (Ed.) United States Department of Agriculture, Forest Service, Washington
Crawley M.J. (1989). Plant life history and the success of weed biological control projects. Pp. 17-26 In: Proceedings of the VII International Symposium on Biological Control of Weeds, E. S. Delfosse (Ed.) Rome, Italy. Istituto Sperimentale per la Patologia Vegetale (MAF).
Cullen J.M. (1989). Current problems in host-specificity screening. Pp. 27-36 In: Proceedings of the VII International Symposium on Biological Control of Weeds, E.S. Delfosse (Ed.) CSIRO Publications, Melbourne.
Cullen J.M. (1995). Predicting effectiveness: fact or fantasy? Pp. 103-109 In: Proceedings of the VIII International Symposium on Biological Control of Weeds, E. S. Delfosse and R. R. Scott (Ed.) Lincoln University, New Zealand, DSIR/CSIRO, Melbourne, Australia.
Davies J.T., Ireson, J.E. and Allen G.R., (2005). The impact of gorse thrips, ryegrass competition, and simulated grazing on gorse seedling performance in a controlled environment. Biological Control 32: 280-286
Duan J.J. and Messing R.H. (1998). Effect of Tetrastichus giffardianus (Hymenoptera: Eulophidae) on nontarget flowerhead-feeding tephritids (Diptera: Tephritidae). Biological Control 27: 1022-1028.
ERMA New Zealand (2000). Pseudaphycus maculipennis for the control of the obscure mealybug (Pseudococcus viburni). Evaluation and Review Report.
FAO (1996). Code of conduct for the import and release of exotic biological control agents. Food and Agriculture Organisation. International Standards for Phytosanitary Measures No 3.
Ferguson C.M., Moeed A., Barratt B.I.P., Hill R.L. and Kean J.M. (2007). BCANZ - Biological Control Agents introduced to New Zealand. http://www.b3nz.org/bcanz
Fowler S.V. and Griffin D. (1995). The effect of multi-species herbivory on shoot growth in gorse Ulex europaeus. Pp. 579-584 In: Proceedings of the VIIIth International Symposium on Biological Control of Weeds, E.S. Delfosse (Ed.) Christchurch, New Zealand
Fowler S.V., Gourlay A.H., Hill R.L. and Withers T. (2003). Safety in New Zealand weed biocontrol: a retrospective analysis of host-specificity testing and the predictability of impacts on non-target plants. Pp. 265�270 in Proceedings of the XI International Symposium on Biological Control of Weeds, Canberra, Australia, 2003, J.M. Cullen, D.T. Briese, D.J. Kriticos, W.M. Lonsdale, L. Morin and J.K. Scott (Ed.).
Fowler S.V., Syrett P. and Hill R.L. (2000). Success and safety in the biological control of environmental weeds in New Zealand. Austral Ecology 25: 553-562.
Froud K.J. and Stevens P.S. (2004). Importation biological control of Heliothrips haemorrhoidalis (Thysanoptera: Thripidae) by Thripobius semiluteus (Hymenoptera: Eulophidae) in New Zealand - a case study of non-target host and environmental risk assessment. Pp. 90-102 In: Assessing host ranges for parasitoids and predators used for classical biological control: a guide to best practice, R. Van Driesche and R. Reardon (Ed.) USDA Forest Service, Morganstown, West Virginia, USA
Froud K.J. and Stevens P.S., (1998). Parasitism of Heliothrips haemorrhoidalis and two non-target thrips by Thripobius semiluteus (Hymenoptera; Eulophidae) in quarantine. Pp. 526-529 In: Pest Management - Future Challenges. Proceedings of the 6th Australasian Applied Entomological Research Conference, M.P. Zalucki, R.A.I. Drew and G.G. White (Ed.) Brisbane Australia.
Fuester R.W., Kenis M., Swan K.S., Kingsley P.C., L�pez-Vaamonde C. and H�rard F. (2001). Host Range of Aphantorhaphopsis samarensis (Diptera: Tachinidae), a larval parasite of the gypsy moth (Lepidoptera: Lymantriidae). Environmental Entomology 25: 332-340.
Gassman A. and Louda S.M. (2001). Rhinocyllus conicus: initial evaluation and subsequent ecological impacts in North America. Pp. 147-183 In: Evaluating indirect ecological effects of biological control, E. Wajnberg, J.K. Scott and P.C. Quimby (Ed.) CABI Publishing, Wallingford, Oxon., UK.
Gassmann A. and Schroeder D. (1995). The search for effective biological control agents in Europe: History and lessons from leafy spurge (Euphorbia esula L.) and cypress spurge (E. cyperissias L.). Biological Control 5: 466-472
Gassmann A., Tosevski I. and Skinner L. (2008). Use of native range surveys to determine the potential host range of arthropod herbivores for biological control of two related weed species, Rhamnus cathartica and Frangula alnus. Biological Control 45: 11-20
Gerard P.J., McNeill M.R., Barratt B.I.P. and Whiteman S.A. (2006). Rationale for release of the irish strain of Microctonus aethiopoides for biocontrol of clover root weevil. New Zealand Plant Protection 59: 285-289.
Godfray H.C.J. (1994). Parasitoids: Behavioural and Evolutionary Ecology. Princeton University Press, Princeton. 473 pp.
Goldson S.L., McNeill M.R. and Proffitt J.R. (2003). Negative effects of strain hybridisation on the biocontrol agent Microctonus aethiopoides. New Zealand Plant Protection 57: 138-142.
Goldson S.L., Phillips C.B., McNeill M.R. and Barlow N.D. (1997). The potential of parasitoid strains in biological control: observations to date on Microctonus spp. intraspecific variation in New Zealand. Agriculture, Ecosystems and Environment 64: 115-124.
Goldson S.L., Proffitt J.R. and Baird D.B. (1998). Establishment and phenology of the parasitoid Microctonus hyperodae (Hymenoptera: Braconidae) in New Zealand. Environmental Entomology 27: 1386-1392.
Goldson S.L., Proffitt J.R. and McNeill M.R. (1990). Seasonal biology and ecology in New Zealand of Microctonus aethiopoides (Hymenoptera: Braconidae), a parasitoid of Sitona spp. (Coleoptera: Curculionidae), with special emphasis on atypical behaviour. Journal of Applied Ecology 27: 703-722.
Goolsby J.A., Van Klinken R.D. and Palmer W.A. (2006). Maximising the contribution of native-range studies towards the identification and prioritisation of weed biocontrol agents. Australian Journal of Entomology 45: 276-286
Gourlay A.H., Wittenberg R., Hill R.L., Spiers A.G. and Fowler S.V. (2000). The biological control programme against Clematis vitalba in New Zealand. Pp. 709-718 In: Proceedings of the X International Symposium on Biological Control of Weeds, N. R. Spencer (Ed.) Bozeman, Montana, USA Montana State University.
Grandgirard J., Hoddle M.S., Petit J.N., Percy D.M. and Roderick G.K. (2006). Pre-introductory risk assessment studies of Gonatocerus ashmeadi (Hymenoptera: Mymaridae) for use as a classical biological control agent against Homalodisca vitripennis (Hemiptera: Cicadellidae) in the Society Islands of French Polynesia. Biocontrol Science & Technology 17: 809-822
Greathead D.J. and Greathead A.H. (1992). Biological control of insect pests by insect parasitoids and predators: the BIOCAT database. Biocontrol News and Information 13: 61N-68N
Grosskopf G., Smith L.A. and Syrett P. (2002). Host range of Cheilosia urbana (Meigen) and Cheilosia psilophthalma (Becker) (Diptera: Syrphidae), candidates for the biological control of invasive alien hawkweeds (Hieracium spp., Asteraceae) in New Zealand. Biological Control 24: 7-19
Gurney W., Crowley P. and Nisbet R. (1992). Locking life-cycles onto seasons: Circle-map models of population dynamics and local adaptation. Journal of Mathematical Biology 30: 251-279.
Hatcher P.E. (1995). Three-way interactions between plant pathogenic fungi, herbivorous insects and their host plants. Biological Reviews 70: 639-694
Hatcher P.E. and Melander B. (2003). Combining physical, cultural and biological methods: prospects for integrated non-chemical weed management strategies. Weed Research 43: 303-322
Hawkins B.A. and Cornell H.V. (1994). Maximum parasitism rates and successful biological control. Science 266: 1886.
Hawkins B.A. and Cornell H.V. (1999). Theoretical approaches to biological control. Cambridge University Press, Cambridge, UK. 412 pp.
Haye T., Goulet H., Mason P.G. and Kuhlmann U. (2005). Does fundamental host range match ecological host range? A retrospective case study of a Lygus plant bug parasitoid. Biological Control 35: 55-67.
Heard T.A. and Van Klinken R.D. (1998). An analysis of test designs for host range determination of insects for biological control of weeds. Pp. 539-546 In: Proceedings of the Sixth Australasian Applied Entomological Research Conference, M.P. Zalucki, R.A.I. Drew and G.G. White (Ed.) Brisbane, University of Queensland.
Hill M.G. (1988). Analysis of the biological control of Mythimna separata (Lepidoptera: Noctuidae) by Apanteles ruficrus (Braconidae: Hymenoptera) in New Zealand. Journal of Applied Ecology 25: 197-208
Hill M.P. and Hulley P.E. (1995). Host-range extension by native parasitoids to weed biocontrol agents introduced to South Africa. Biological Control 5: 297-302.
Hill R.L. (1977). Parasite helps control armyworm. New Zealand Journal of Agriculture 134: 21-23
Hill R.L. (1999). Minimising uncertainty - in support of no-choice tests. Pp. 1-10 In: Host specificity testing in Australasia: towards improved assays for biological control, W.T.M., L. Barton Browne and J. N. Stanley (Ed.) CRC for Tropical Pest Management, Brisbane, Australia.
Hill R.L. (2008). Application to release from containment a beetle, Lema obscura F. (Chrysomelidae), for the biological control of the weed tradescantia (Tradescantia fluminensis). http://www.ermanz.govt.nz/appfiles/execsumm/pdf/NOR07001-002.pdf
Hill R.L. and Gourlay A.H. (2002). Host-range testing, introduction, and establishment of Cydia succedana (Lepidoptera: Tortricidae) for biological control of gorse, Ulex europaeus L., in New Zealand. Biological Control 25: 173-186
Hill R.L. and O'Donnell D.J. (1991). Reproductive isolation between Tetranychus lintearius and two related mites, T. urticae and T. turkestani (Acarina: Tetranychidae). Experimental and Applied Acarology 11: 241-251
Hill R.L. and Sandrey R.A. (1986). The costs and benefits of gorse Proceedings of the New Zealand Weed and Pest Control Conference 39: 70-73
Hill R.L., Cumber R.A. and Allan D.J. (1985). Parasitoids introduced to control larvae of the Noctuidae (Lepidoptera) in New Zealand (1968-1978). DSIR Entomology Division, 24 p.
Hill R.L., Gourlay, A.H. and Winks, C.J. (1993). Choosing gorse spider mite strains to improve establishment in different climates. In: Proceedings of the 6th Australasian Grasslands Invertebrate Ecology Conference, R.A Prestidge (Ed.). AgResearch, Hamilton, New Zealand.
Hill R.L., Markin G.P., Gourlay A.H., Fowler S.V. and Yoshioka E. (2001). Host range, release, and establishment of Sericothrips staphylinus Haliday (Thysanoptera: Thripidae) as a biological control agent for gorse, Ulex europaeus L. (Fabaceae), in New Zealand and Hawaii. Biological Control 21: 63-74
Hill R.L., Wittenberg R. and Gourlay A.H. (2001). Biology and host range of Phytomyza vitalbae and its establishment for the biological control of Clematis vitalba in New Zealand. Biocontrol Science and Technology 11: 459-473
Hoddle M. (2004). Restoring balance using exotic species to control invasive exotic species. Conservation Biology 18: 38-49
Hokkanen H.M.T. and Pimentel D. (1989). New associations in biological control: Theory and practice. Canadian Entomologist 121: 829-840
Holt R.D. and Hochberg M.E. (2001). Indirect interactions, community modules and biological control: a theoretical perspective. Pp. 13-37 In: Evaluating indirect ecological effects of biological control, E. Wajnberg, J. K. Scott and P. C. Quimby (Ed.) CABI Publishing, Wallingford, Oxon., UK
Hopper K.R. and Wajnberg E. (2006). The risks of interbreeding and methods for determination. In press In: Environmental Impact of Arthropod Biological Control: Methods and Risk Assessment, U. Kuhlmann, F. Bigler and D. Babendreier (Ed.) CABI Bioscience, Delemont, Switzerland.
Hufbauer R.A. and Roderick G.K. (2005). Microevolution in biological control: Mechanisms, patterns, and processes. Biological Control 35: 227-239.
Jarvis P.J., Fowler S.V., Paynter Q. and Syrett P. (2006). Predicting the economic benefits and costs of introducing new biological control agents for Scotch broom Cytisus scoparius into New Zealand. Biological Control 39: 135-146
Julien M.H., Skarratt B. and Maywald G.F. (1995). Potential geographical distribution of alligator weed and its biological control by Agasicles hygrophila. Journal of Aquatic Plant Management 2: 55-60
Julien, M. and Griffiths M.W. (1999). Biological Control of Weeds. A World Catalogue of Agents and their Target Weeds. CABI Publishing, Wallingford, UK. 223 p.
Kay N. and Hill R.L. (in press). The disintegration of the Scrophulariaceae and the biological control of Buddleja davidii. In: Proceedings of the XII International Symposium on Biological Control of Weeds, (Ed.) Montpellier, France.
Kean J.M. and Barlow N.D. (2000). Can host-parasitoid metapopulations explain successful biological control? Ecology 81: 2188-2197.
Kean J.M. and Kumarasinghe L. (2007). Predicting the seasonal phenology of fall webworm (Hyphantria cunea) in New Zealand. New Zealand Plant Protection 60: 279-285.
Kindlmann P. and Dixon A.F.G. (1999). Generation time ratios - determinants of prey abundance in insect predator-prey interactions. Biological Control 16: 133-138.
Kitt J.T. and Keller M.A. (1998). Host selection by Aphidius rosae Haliday (Hym., Braconidae) with respect to assessment of host specificity in biological control. Journal of Applied Entomology 122: 57-63
Kriticos D.J. and Randall R.P. (2001). A comparison of systems to analyse potential weed distributions. Pp. 61-79 in: Groves R.H., Panetta F.D. and Virtue J.G. ed. Weed Risk Assessment. CSIRO Publishing, Melbourne, Australia.
Kriticos D.J., Brown J.R., Radford I.D. and Nicholas M. (1999). Plant population ecology and biological control: Acacia nilotica as a case study. Biological Control 16
Krombein K.V., Hurd P.D., Smith D.R. and Burks B.D. (1979). Catalog of Hymenoptera in America North of Mexico. Smithsonian Institution Press, Washington, DC, USA
Kuhlmann U., Mason P.G., Hinz H.L., Blossey B., De Clerck-Floate R.A., Dosdall L.M., McCaffrey J.P., Schwarzlaender M., Olfert O., Brodeur J., Gassmann A., McClay A.S. and Wiedenmann R.N. (2006). Avoiding conflicts between insect and weed biological control: selection of non-target species to assess host specificity of cabbage seedpod weevil parasitoids. Journal of Applied Entomology 130: 129-141
Kuhlmann U., Schaffner U. and Mason P.G. (2005). Selection of non-target species for host specificity testing of entomophagous biological control agents. Pp. 566-583 In: Second International Symposium on Biological Control of Arthropods, Davos, Switzerland, 12-16 September, 2005, M.S. Hoddle (Ed.) United States Department of Agriculture, Forest Service, Washington.
Lockwood J.A. (2000). Nontarget effects of biological control: what are we trying to miss? Pp. 15-30 In: Nontarget effects of biological control introductions, P.A. Follett and J.J. Duan (Ed.) Kluwer Academic Publishers, Norwell, Massachusetts, USA.
Louda S.M., Pemberton R.W., Johnson M.T. and Follett P.A. (2003). Nontarget effects - the Achilles' heel of biological control? Retrospective analyses to reduce risk associated with biocontrol introductions. Annual Review of Entomology 48: 365-396.
Louda S.M., Rand T.A., Russell F.L. and Arnett A.E. (2005). Assessment of ecological risks in weed biocontrol: Input from retrospective ecological analyses. Biological Control 35: 253-264.
Lynch L.D. and Thomas M.B. (2000). Nontarget effects in the biocontrol of insects with insects, nematodes and microbial agents: the evidence. Biocontrol News and Information 21: 117N-130N
Malone L.A., Todd J.H., Burgess E.P.J., Walter C., Wagner A. and Barratt B.I.P. (2010). Developing risk hypotheses and selecting species for assessing non-target impacts of GM trees with novel traits: the case of altered-lignin pine trees. Environmental Biosafety Research 9: 181-198.
Marohasy J. (1998). The design and interpretation of host-specificity tests for weed biological control with particular reference to insect behaviour. Biocontrol News and Information 19: 13-20.
Mason P.G., Broadbent A.B., Whistlecraft J.W. and Gillespie D.R. (2011). Interpreting the host range of Peristenus digoneutis and Peristenus relictus (Hymenoptera: Braconidae) biological control agents of Lygus spp. (Hemiptera: Miridae) in North America. Biological Control 57: 94-102.
McClay A.S. and Balciunas J.K. (2005). The role of pre-release efficacy assessment in selecting classical biological control agents for weeds - applying the Anna Karenina principle. Biological Control 35: 197-207
McEvoy P.B. and Coombs E.M. (1999). Biological control of plant invaders: regional patterns, field experiment and structured population models. Ecological Applications 9: 387-401
McFadyen R.E.C. (1998). Biological control of weeds. Annual Review of Entomology 43: 369-393.
McNeill M.R., Ferguson C.M., Bixley A.S. and Barratt B.I.P. (2009). Development of Microctonus aethiopoides Loan through multiple generations of novel weevil hosts in the laboratory. Pp. 617-618 in: Proceedings of the 3rd International Symposium on Biological Control of Arthropods, P.G. Mason, D.R. Gillespie and C. Vincent (Ed.) Christchurch, New Zealand.
Messing R.H., Roitberg B.D. and Brodeur J. (2006). Measuring and predicting indirect impacts of biological control, competition, displacement and secondary interactions. Pp. 64-77 In: Environmental impact of invertebrates for biological control of arthropods - methods and risk assessment, F. Bigler, D. Babendreier and U. Kuhlmann (Ed.) CABI Publishing, Wallingford, UK
Mills N.J. (2006). Accounting for differential success in the biological control of homopteran and lepidopteran pests. New Zealand Journal of Ecology 30: 61-72.
Mills N.J. and Getz W.M. (1996). Modelling the biological control of insect pests: a review of host-parasitoid models. Ecological Modelling 92: 121-143
Mills N.J. and Gutierrez A.P. (1999). Biological contro of insects: a tritrophic perspsective. Pp. 89-102 In: Theoretical approaches to biological control, B. A. Hawkins and H. V. Cornell (Ed.) Cambridge University Press, Cambridge, UK
Mills N.J. and Kean J.M. (2010). Modelling, behavioural studies and molecular approaches: Methodological contributions to biological control success. Biological Control (in press).
Morin L. and Edwards P.B. (2006). Selection of biological control agents for bridal creeper: a retrospective review. Australian Journal of Entomology 45: 287-291
Morin L., Evans K.J. and Sheppard A.W. (2006). Selection of pathogen agents in weed biological control: critical issues and peculiarities in relation to arthropod agents. Australian Journal of Entomology 45: 349-365
Morin L., Hill R.L. and Matayoshi S. (1997). Hawaii's successful biological control strategy for mist flower (Ageratina riparia) - can it be transferred to New Zealand? Biocontrol News and Information 18: 77-88
Munro V.M.W. and Henderson I.F. (2002). Nontarget effect of entomophagous biocontrol: shared parasitism between native lepidopteran parasitoids and the biocontrol agent Trigonospila brevifacies (Diptera: Tachinidae) in forest habitats. Environmental Entomology 32: 388-396.
Paynter Q., Waipara N.W., Peterson P.G., Hona S.R., Fowler S.V., Gianotti A.F. and Wilkie P. (2006). The impact of two introduced biocontrol agents, Phytomyza vitalbae and Phoma clematidina, on Clematis vitalba in New Zealand. Biological Control 36: 350-357
Paynter Q.E., Fowler S.V., Gourlay A.H., Haines M.L., Harman H.M., Hona S.R., Peterson P.G., Smith L.A., Wilson-Davey, J.R.A., Winks, C.J. and Withers T.M. (2004). Safety in New Zealand Weed Biocontrol: A nationwide survey for impacts on non-target plants. New Zealand Plant Protection 57: 102-107
Pearson D.E. and Callaway R.M. (2005). Indirect nontarget effects of host-specific biological control agents: Implications for biological control. Biological Control 35: 288-298.
Pemberton R.W. (2000). Predictable risk to native plants in weed biological control. Oecologia 125: 489-494.
Pennacchio F. and Strand M.R. (2006). Evolution of developmental strategies in parasitic Hymenoptera. Annual Review of Entomology 51: 233-258
Phillips C.B., Proffitt J.R. and Goldson S.L. (1998). Potential to enhance the efficacy of Microctonus hyperodae Loan. Proceedings of the New Zealand Plant Protection Conference 51: 16-22.
Powell J.A. and Logan J.A. (2005). Insect seasonality: circle map analysis of temperature-driven life cycles. Theoretical Population Biology 67: 161-179.
Price P.W. (2000). Host plant resource quality, insect herbivores and biocontrol. Pp. 583-590 In: Proceedings of the Xth International Symposium on Biological Control of Weeds, N.R. Spencer (Ed.) Montana State University, Bozeman Montana, USA
Raghu S. and Van Klinken R.D. (2006). Refining the ecological basis for agent selection in weed biological control. Australian Journal of Entomology 45: 251-252
Raghu S., Dhileepan K. and Scanlan J.C. (2007). Predicting risk and benefit a priori in biological control of invasive plant species: A systems modelling approach. Ecological Modelling 208: 247-262
Raghu S., Wilson J.R. and Dhileepan K. (2006). Refining the process of agent selection through understanding plant demography and plant response to herbivory. Australian Journal of Entomology 45: 308-316
Romeis J., Babendreier D., Wackers F.L. and Shanower T.G. (2005). Habitat and plant specificity of Trichogramma egg parasitoids - underlying mechanisms and implications. Basic and Applied Ecology 6: 215-236
Rutledge C.E. and Wiedenmann R.N. (1999). Habitat preferences of three congeneric braconid parasitoids: implications for host-range testing in biological control. Biological Control 16: 144-154
Sands D.P.A. and Van Driesche R.G. (2004). Using the scientific literature to estimate the host range of a biological control agent. Pp. 15-23 In: Assessing host ranges for parasitoids and predators used for classical biological control: a guide to best practice, R.G. Van Driesche and R. Reardon (Ed.) USDA Forest Service, Morgantown, West Virginia.
Sheppard A., Haines M.L. and Thomann T. (2006). Native-range research assists risk analysis for non-targets in weed biological control: the cautionary tale of the broom seed beetle. Australian Journal of Entomology 45: 292-297
Sheppard A.W. (1999). Which test? A mini review of test usage in host specificity testing. Pp. 60-69 In: Host specificity testing in Australasia: towards improved assays for biological control, T.M. Withers, L. Barton-Browne and J. Stanley (Ed.) Scientific Publishing, Department of Natural Resources, Brisbane.
Sheppard A.W. (2003). Prioritising agents based on predicted efficacy: beyond the lottery approach. Pp. 11-21 In: Improving the selection, testing and evaluation of weed biocontrol agents, H. Spafford-Jacobs and D. T. Briese (Ed.) Cooperative Research Centre for Australian Weed Management, Adelaide, Australia
Simberloff D. and Stiling P. (1996). Risks of species introduced for biological control. Biological Conservation 78: 185-192.
Spafford-Jacob H. and Briese D.T. (2003). Improving the selection, testing and evaluation of weed biocontrol agents. CRC for Australian Weed Management Technical Series no. 7, Adelaide, Australia.
Spradberry P. (1970). Host finding by Rhyssa persuasoria L. an ichneumon parasite of siricid woodwasps. Animal Behaviour 18: 103-114
Standish R.J., Robertson A.W. and Williams P.A. (2001). The impact of an invasive weed Tradescantia fluminensis on native forest regeneration. Journal of Applied Ecology 38: 1253-1263
Stufkens M.W., Farrell J.A. and Popay A.J. (1994). Quarantine host range tests on two exotic parasitoids imported for aphid control. Pp. 149-153 In: Proceedings of the 47th New Zealand Plant Protection Conference.
Sutherst R.W. and Maywald G.F. (1985). A computerised system for matching climates in ecology. Agriculture, Ecosystems and Environment 13: 281-299.
Sutherst R.W., Maywald G.F. and Kriticos D.J. (2007). CLIMEX version 3. CD and User's Guide. Hearne Scientific Software, Melbourne, Australia.
Syrett P. and Emberson R.M. (1997). The natural host range of beetle species feeding on broom Cytisus scoparius (L.) Link (Fabaceae) in south-west Europe. Biocontrol Science and Technology 7: 309-326
Thomas M. and Waage J.K. (1996). Integration of biological control and host plant resistance breeding. A scientific and literature review. Technical Centre for Agricultural and Rural Cooperation (CTA), 99 pp.
Thompson W.R. and Simmonds F.J. (1964-1965). A Catalogue of the Parasites and Predators of Insect Pests. Commonwealth Agricultural Bureaux, Bucks, United Kingdom.
Todd J.H., Barratt B.I.P., Tooman L., Beggs J.R. and Malone L.A. (2015). Selecting non-target species for risk assessment of entomophagous biological control agents: Evaluation of the PRONTI decision-support tool. Biological Control 80: 77-88.
Todd J.H., Ramankutty P., Barraclough E.I. and Malone L.A. (2008). A screening method for prioritizing non-target invertebrates for improved biosafety testing of transgenic crops. Environmental Biosafety Research 7: 35-56.
Turlings T.C.J., Tumlinson J.H., Heath R.R., Proveaux, A.T. and Doolittle R.E. (1991). Isolation and identification of allelochemicals that attract the larval parasitoid, Cotesia marginiventris (Cresson), to the microhabitat of one of its hosts. Journal of Chemical Ecology 17: 2235-2251
van Driesche R. (2004). Predicting host ranges of parasitoids and predacious insects - what are the issues? Pp. 1-3 In: Assessing host ranges for parasitoids and predators used for classical biological control: a guide to best practice, R. Van Driesche and R. Reardon (Ed.) USDA Forest Service, Morgantown, West Virginia.
van Driesche R. and Reardon R. (2004). Assessing host ranges for parasitoids and predators used for classical biological control: a guide to best practice. Pp. 243. USDA Forest Service, Morgantown, West Virginia.
van Driesche R.G. and Hoddle M. (1997). Should arthropod parasitoids and predators be subject to host range testing when used as biological control agents? Agriculture and Human Values 14: 211-226.
van Driesche R.G. and Hoddle M.S. (2000). Classical arthropod biological control: assessing success, step by step. Pp. 39-75 In: Biological control: Measures of success, G.M. Gurr and S.D. Wratten (Ed.) Kluwer Academic Publishers, Dordrecht, The Netherlands
van Driesche R.G. and Murray T.J. (2004). Overview of testing schemes and designs used to estimate host ranges. Pp. 68-89 In: Assessing host ranges for parasitoids and predators used for classical biological control: a guide to best practice, R.G. Van Driesche and R. Reardon (Ed.) USDA Forest Service, Morgantown, West Virginia.
van Klinken R.D. (2006). Biological control of Parkinsonia aculeata: what are we trying to achieve? Australian Journal of Entomology 45: 268-271
van Klinken R.D. and Raghu S. (2006). A scientific approach to agent selection. Australian Journal of Entomology 45: 253-258
van Lenteren J.C. (2006). The internet book of biological control http://www.unipa.it/iobc/downlaod/IOBC%20InternetBookBiCoVersion4October2006.pdf
van Lenteren J.C., Bale J., Bigler F., Hokkanen H.M.T. and Loomans A.J.M. (2006). Assessing risks of releasing exotic biological control agents of arthropod pests. Annual Review of Entomology 51: 609-634.
van Lenteren J.C., Cock M.J.W., Hoffmeister T.S. and Sands D.P.A. (2006). Host specificity in arthropod biological control, methods for testing and interpreting the data. Pp. 38-63. CAB Publishing, Delemont.
van Veen F.J.F., Morris R.J. and Godfray H.C.J. (2006). Apparent competition, quantitative food webs, and the structure of phytophagous insect communities. Annual Review of Entomology 51: 187-208
Vet L.E.M. and Dicke M. (1992). Ecology of infochemical use by natural enemies in a tritrophic context. Annual Review of Entomology 37: 141-172
Vet L.E.M. and Godfray H.C.J. (2008). Multitrophic interactions and parasitoids behavioural ecology. Pp. 231-253 In: Behavioural ecology of insect parasitoids: from theoretical approaches to field application, E. Wajnberg, C. Bernstein and J.J.M. Van Alphen (Ed.) Blackwell Publishing, Oxford, UK
Vink C.J. and Phillips C.B. (2007). First record of Sitona discoideus Gyllenhal 1834 (Coleoptera: Curculionidae) on Norfolk Island. New Zealand Journal of Zoology 34: 283-287.
Vink C.J., Phillips C.B., Mitchell A.D., Winder L.M. and Cane R.P. (2003). Genetic variation in Microctonus aethiopoides (Hymenoptera: Braconidae). Biological Control 28: 251-264
Vinson S.B. (1998). The general host selection behavior of parasitoid Hymenoptera and a comparison of initial strategies utilized by larvaphagous and oophagous species. Biological Control 11: 79-96
Wapshere A.J. (1974). A strategy for evaluating the safety of organisms for biological weed control. Annals of Applied Biology 77: 201-211.
Watson M.C. (2007). Buddleia weevil blooms. What's New in Biological Control of Weeds 41: 11.
Wearing C.H. (1979). Integrated control of apple pests in New Zealand 10. Population dynamics of codling moth in Nelson. New Zealand Journal of Zoology 6: 165-199
Wearing C.H. and Charles J.G. (1989). Cydia pomonella (L.), codling moth (Lepidoptera: Tortricidae). Pp. 161-169 In: A review of biological control of insect pests and weeds in New Zealand 1874 to 1987, P.J. Cameron, R.L. Hill, J. Bain and W.P. Thomas (Ed.) CAB International Institute of Biological Control 10, CAB International, Wallingford, UK
Willis A.J. and Memmott J. (2005). The potential for indirect effects between a weed, one of its biocontrol agents and native herbivores: A food web approach. Biological Control 35: 299-306.
Withers T.M. and Barton-Browne L. (2004). Behavioral and physiological processes affecting the outcome of host range testing. Pp. 40-55 In: Assessing host ranges for parasitoids and predators used for classical biological control: a guide to best practice, R.G. Van Driesche and R. Reardon (Ed.) USDA Forest Service, Morgantown, West Virginia.
Withers T.M., Allen G.R. and Reid C.A.M. (2015). Selecting potential non-target species for host range testing of Eadya paropsidis. New Zealand Plant Protection 68: 179-186.
Withers T.M., Hill R.L., Paynter Q., Fowler S.V. and Gourlay A.H. (2008). Post-release investigations into the field host range of the gorse pod moth Cydia succedana Denis & Schiffermuller (Lepidoptera : Tortricidae) in New Zealand. New Zealand Entomologist 31: 67-76
 Previous page
Previous page
Background information Next page
Next page
Applying to import into containment