
Selecting biological control agents
Predicting direct non-target impacts
Modelling non-target impacts
Attack rate and population impact
The success of a BCA is usually estimated by measuring the proportion of the pest or weed population that is attacked in the field. Similarly, nontarget impacts are usually quantified in terms of the proportion of nontarget individuals attacked (Mills and Kean 2010). However, this is misleading since host population size may or may not be significantly affected by a BCA, irrespective of the proportion attacked (Barratt et al. 2010). Relative population size with and without BCA attack will almost always be the most relevant measure of BCA impact, so the relationship between attack rate and population size is fundamental to assessing impact.
Kean and Barlow (2000) and Barlow et al. (2004) were the first to use models to explicitly explore the relationship between BCA attack rate and host population suppression, in the contexts of target and non-target impacts, respectively. These models have been advanced by developing an overarching framework which encompasses any enemy-host life cycle, relates the previous models to each other, and suggests a hypothesis to explain the empirical relationship between maximum attack rate and the probability of successful biological control noted by Hawkins and Cornell (1994).
Estimation of population impact from a given attack rate, and the attack rate leading to host extinction, requires knowledge of the host life cycle and intrinsic rate of increase; these are basic parameters that may be estimated relatively easily. For deliberate introductions, indicative attack rates may be estimated from pre-release host range testing experiments. Alternatively, the models are simple enough that decision-makers can easily simulate "what if?" scenarios to explore the implications of uncertainty in the parameters. An interactive web page is available for this purpose.
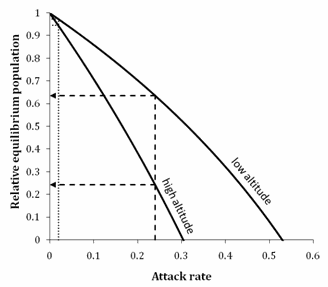
Relationship between attack rate and impact for low and high altitude native weevil populations in Otago.
The figure shows an application of this model to native weevils under nontarget attack by the parasitoid Microctonus aethiopoides in Otago tussock grasslands. The intrinsic rate of increase of these weevil populations declines with altitude (Barlow et al. 2004). While neither upland nor lowland populations are likely to be significantly reduced by the 2% average attack rate observed (dotted line), both would be substantially reduced by the 24% maximum attack rate observed (Barratt et al. 2007), should this be sustained (dashed line). Under the latter scenario, the model suggests that lowland populations would be reduced to 64% of their natural density and upland populations to 24%. The model also suggests that lowland populations will be driven extinct by sustained attack rates of greater than 53% and upland populations by greater than 30%. The ability to estimate population impact from attack rate will be of enormous benefit to the EPA's and DOC's reviews of proposals for future proposed BCA introductions.
Future research should emphasise validation of the models using meta-analysis of published results from the literature, and post-release studies of biological control introductions such as those being conducted in projects being carried out in the research programme Better Border Biosecurity [http://www.b3nz.org].
References
Barlow N.D., Barratt B.I.P., Ferguson C.M. and Barron M.C. (2004). Using models to estimate parasitoid impacts on non-target host abundance. Environmental Entomology 33: 941-948.
Barratt B.I.P., Ferguson C.M., Bixley A.S., Crook K.E., Barton D.M. and Johnstone P.D. (2007). Field parasitism of nontarget weevil species (Coleoptera : Curculionidae) by the introduced biological control agent Microctonus aethiopoides Loan (Hymenoptera : Braconidae) over an altitude gradient. Environmental Entomology 36: 826-839
Barratt B.I.P., Howarth F., Withers T.M., Kean J.M. and Ridley G. (2010). Progress in risk assessment for classical biological control. Biological Control 52: (3) 245-254.
Hawkins B.A. and Cornell H.V. (1994). Maximum parasitism rates and successful biological control. Science 266: 1886.
Kean J.M. and Barlow N.D. (2000). Can host-parasitoid metapopulations explain successful biological control? Ecology 81: 2188-2197.
Mills N.J. and Kean J.M. (2010). Modelling, behavioural studies and molecular approaches: Methodological contributions to biological control success. Biological Control (in press).
Modelling non-target impacts | Interactive model for predicting impact |
|---|